

Comprehensive identification of SWI/SNF complex subunits underpins deep eukaryotic ancestry and reveals new plant components
- PMID: 35668117
- PMCID: PMC9170682
- DOI: 10.1038/s42003-022-03490-x
Comprehensive identification of SWI/SNF complex subunits underpins deep eukaryotic ancestry and reveals new plant components
Abstract
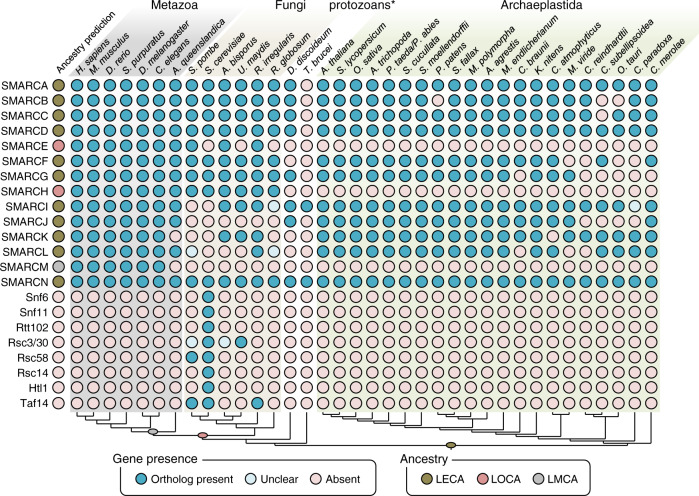
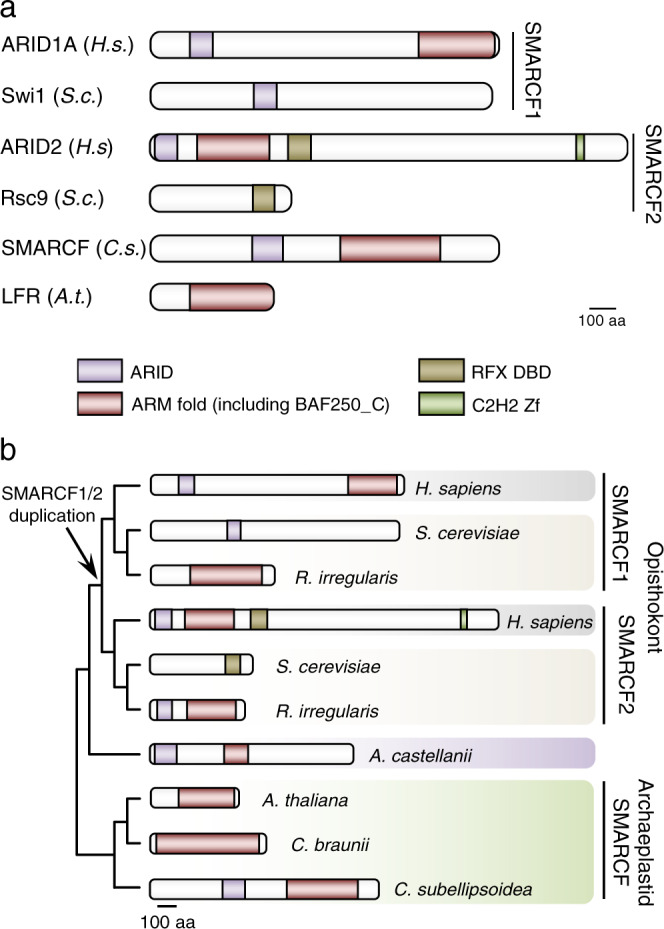
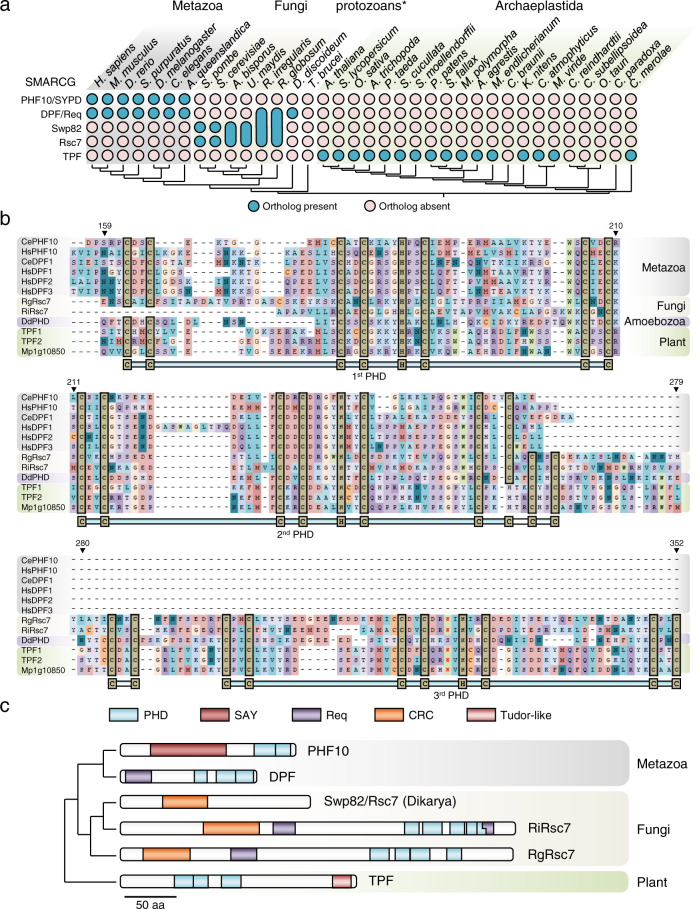
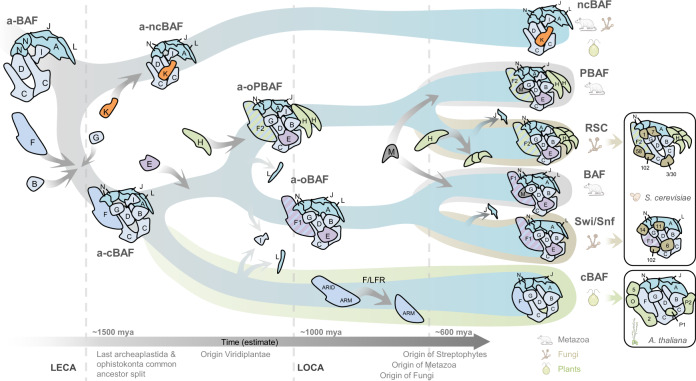
Over millions of years, eukaryotes evolved from unicellular to multicellular organisms with increasingly complex genomes and sophisticated gene expression networks. Consequently, chromatin regulators evolved to support this increased complexity. The ATP-dependent chromatin remodelers of the SWI/SNF family are multiprotein complexes that modulate nucleosome positioning and appear under different configurations, which perform distinct functions. While the composition, architecture, and activity of these subclasses are well understood in a limited number of fungal and animal model organisms, the lack of comprehensive information in other eukaryotic organisms precludes the identification of a reliable evolutionary model of SWI/SNF complexes. Here, we performed a systematic analysis using 36 species from animal, fungal, and plant lineages to assess the conservation of known SWI/SNF subunits across eukaryotes. We identified evolutionary relationships that allowed us to propose the composition of a hypothetical ancestral SWI/SNF complex in the last eukaryotic common ancestor. This last common ancestor appears to have undergone several rounds of lineage-specific subunit gains and losses, shaping the current conformation of the known subclasses in animals and fungi. In addition, our results unravel a plant SWI/SNF complex, reminiscent of the animal BAF subclass, which incorporates a set of plant-specific subunits of still unknown function.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
