

Complete genome sequences and genomic characterization of five plasmids harbored by environmentally persistent Cronobacter sakazakii strains ST83 H322 and ST64 GK1025B obtained from powdered infant formula manufacturing facilities
- PMID: 35668537
- PMCID: PMC9169379
- DOI: 10.1186/s13099-022-00500-5
Complete genome sequences and genomic characterization of five plasmids harbored by environmentally persistent Cronobacter sakazakii strains ST83 H322 and ST64 GK1025B obtained from powdered infant formula manufacturing facilities
Abstract
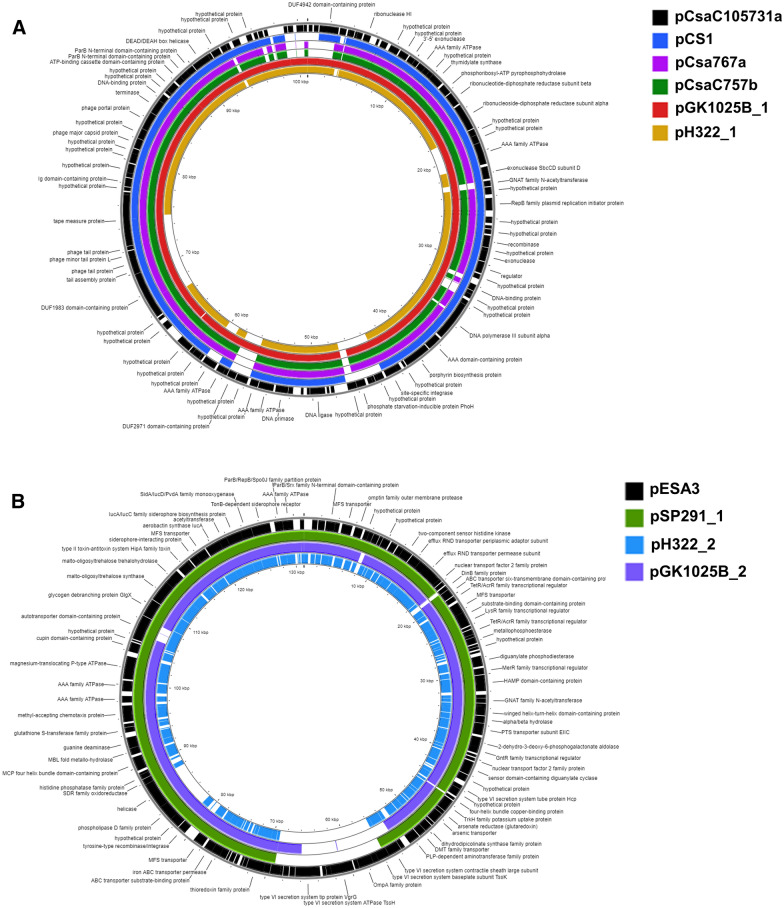
Background: Cronobacter sakazakii is a foodborne pathogen that causes septicemia, meningitis, and necrotizing enterocolitis in neonates and infants. The current research details the full genome sequences of two extremely persistent C. sakazakii strains (H322 and GK1025B) isolated from powdered infant formula (PIF) manufacturing settings. In addition, the genetic attributes associated with five plasmids, pH322_1, pH322_2, pGK1025B_1, pGK1025B_2, and pGK1025B_3 are described.
Materials and methods: Using PacBio single-molecule real-time (SMRT®) sequencing technology, whole genome sequence (WGS) assemblies of C. sakazakii H322 [Sequence type (ST)83, clonal complex [CC] 83) and GK1025B (ST64, CC64) were generated. Plasmids, also sequenced, were aligned with phylogenetically related episomes to determine, and identify conserved and missing genomic regions.
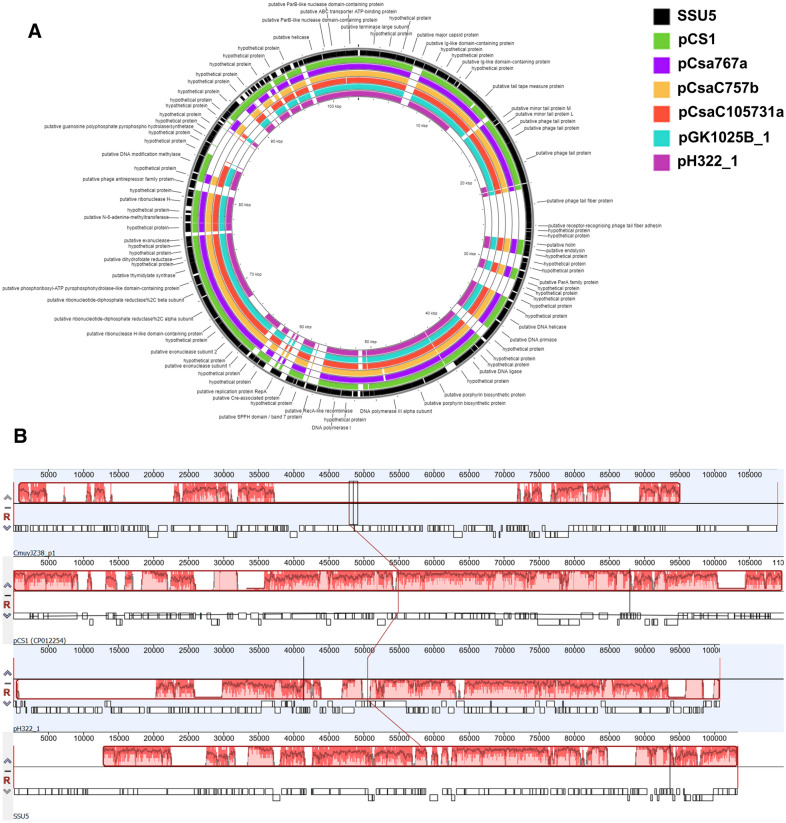
Results: A truncated ~ 13 Kbp type 6 secretion system (T6SS) gene cluster harbored on virulence plasmids pH322_2 and pGK1025B_2, and a second large deletion (~ 6 Kbp) on pH322_2, which included genes for a tyrosine-type recombinase/integrase, a hypothetical protein, and a phospholipase D was identified. Within the T6SS of pH322_2 and pGK1025B_2, an arsenic resistance operon was identified which is in common with that of plasmids pSP291_1 and pESA3. In addition, PHASTER analysis identified an intact 96.9 Kbp Salmonella SSU5 prophage gene cluster in pH322_1 and pGK1025B_1 and showed that these two plasmids were phylogenetically related to C. sakazakii plasmids: pCS1, pCsa767a, pCsaC757b, pCsaC105731a. Plasmid pGK1025B_3 was identified as a novel conjugative Cronobacter plasmid. Furthermore, WGS analysis identified a ~ 16.4 Kbp type 4 secretion system gene cluster harbored on pGK1025B_3, which contained a phospholipase D gene, a key virulence factor in several host-pathogen diseases.
Conclusion: These data provide high resolution information on C. sakazakii genomes and emphasizes the need for furthering surveillance studies to link genotype to phenotype of strains from previous investigations. These results provide baseline data necessary for future in-depth investigations of C. sakazakii that colonize PIF manufacturing facility settings and genomic analyses of these two C. sakazakii strains and five associated plasmids will contribute to a better understanding of this pathogen's survival and persistence within various "built environments" like PIF manufacturing facilities.
Keywords: Built environment; Complete genomes; Cronobacter sakazakii; PHASTER; Phage-plasmids; Plasmids; Whole genome sequencing.
© 2022. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures


References
-
- Jang H, Gopinath GR, Eshwar A, Srikumar S, Nguyen S, Gangiredla J, Patel IR, Finkelstein SB, Negrete F, Woo J, Lee Y, Fanning S, Stephan R, Tall BD, Lehner A. The secretion of toxins and other exoproteins of Cronobacter: role in virulence, adaption, and persistence. Microorganisms. 2020;8(2):229. doi: 10.3390/microorganisms8020229. - DOI - PMC - PubMed
-
- Holý O, Petrželová J, Hanulík V, Chromá M, Matoušková M, Forsythe SJ. Epidemiology of Cronobacter spp. isolates from patients admitted to the Olomouc University Hospital (Czech Republic) Epidemiol Mikrobiol Imunol. 2014;63:69–72. - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
