

WUSCHEL-related homeobox genes cooperate with cytokinin to promote bulbil formation in Lilium lancifolium
- PMID: 35670734
- PMCID: PMC9773970
- DOI: 10.1093/plphys/kiac259
WUSCHEL-related homeobox genes cooperate with cytokinin to promote bulbil formation in Lilium lancifolium
Abstract
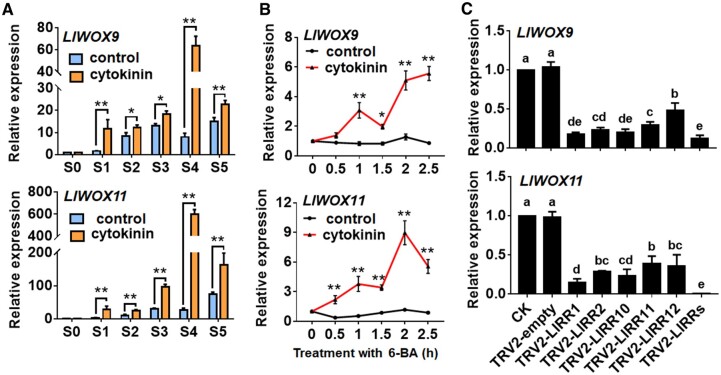
The bulbil is an important vegetative reproductive organ in triploid tiger lily (Lilium lancifolium). Based on our previously obtained transcriptome data, we screened two WUSCHEL-related homeobox (WOX) genes closely related to bulbil formation, LlWOX9 and LlWOX11. However, the biological functions and regulatory mechanisms of LlWOX9 and LlWOX11 are unclear. In this study, we cloned the full-length coding sequences of LlWOX9 and LlWOX11. Transgenic Arabidopsis (Arabidopsis thaliana) showed increased branch numbers, and the overexpression of LlWOX9 and LlWOX11 in stem segments promoted bulbil formation, while the silencing of LlWOX9 and LlWOX11 inhibited bulbil formation, indicating that LlWOX9 and LlWOX11 are positive regulators of bulbil formation. Cytokinin type-B response regulators could bind to the promoters of LlWOX9 and LlWOX11 and promote their transcription. LlWOX11 could enhance cytokinin pathway signaling by inhibiting the transcription of type-A LlRR9. Our study enriches the understanding of the regulation of plant development by the WOX gene family and lays a foundation for further research on the molecular mechanism of bulbil formation in lily.
© American Society of Plant Biologists 2022. All rights reserved. For permissions, please email: journals.permissions@oup.com.
Figures








References
-
- Abraham-Juárez MJ, Martínez-Hernández A, Leyva-González MA, Herrera-Estrella L, Simpson J (2010) Class I KNOX genes are associated with organogenesis during bulbil formation in Agave tequilana. J Exp Bot 61: 4055–4067 - PubMed
-
- Bach A, Sochacki D (2012) Propagation of ornamental geophytes: physiology and management. InKamenetsky R, Okubo H, eds, Ornamental Geophytes: From Basic Science to Sustainable Production. CRC Press, Boca Raton, FL, pp 261–286
-
- Bell AD, Bryan A (2008) Plant Form: An Illustrated Guide to Flowering Plant Morphology. Timber Press, Portland, Oregon
-
- Breuninger H, Rikirsch E, Hermann M, Ueda M, Laux T (2008) Differential expression of WOX genes mediates apical-basal axis formation in the Arabidopsis embryo. Dev Cell 14: 867–876 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
