Murine cytomegalovirus promotes renal allograft inflammation via Th1/17 cells and IL-17A
- PMID: 35671112
- PMCID: PMC9547825
- DOI: 10.1111/ajt.17116
Murine cytomegalovirus promotes renal allograft inflammation via Th1/17 cells and IL-17A
Abstract
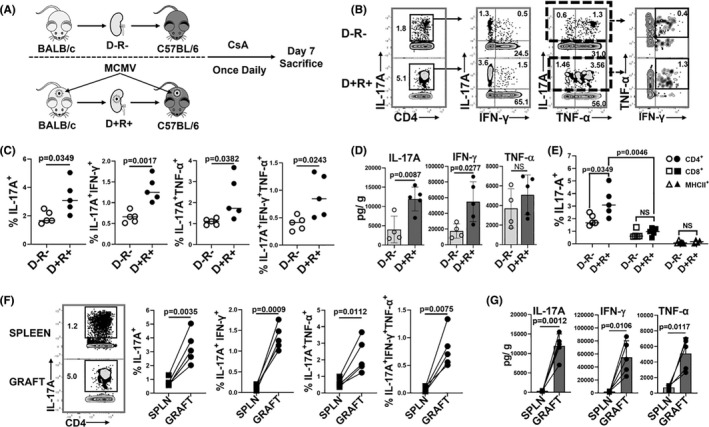
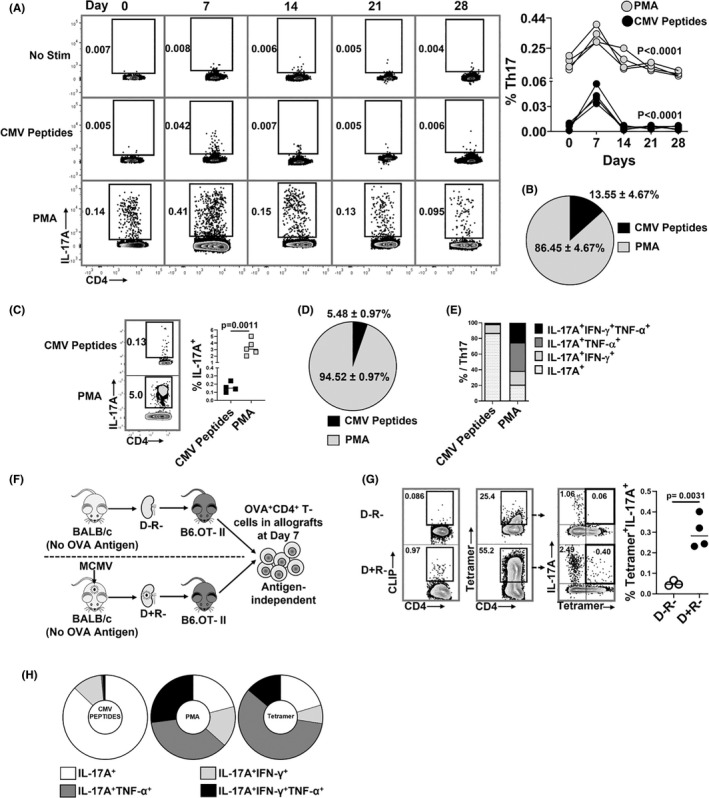
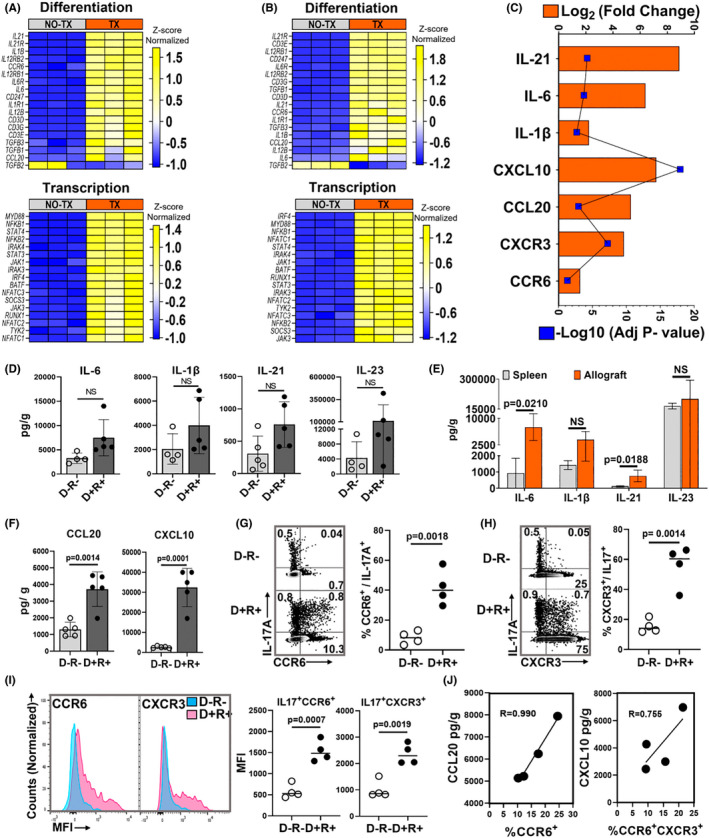
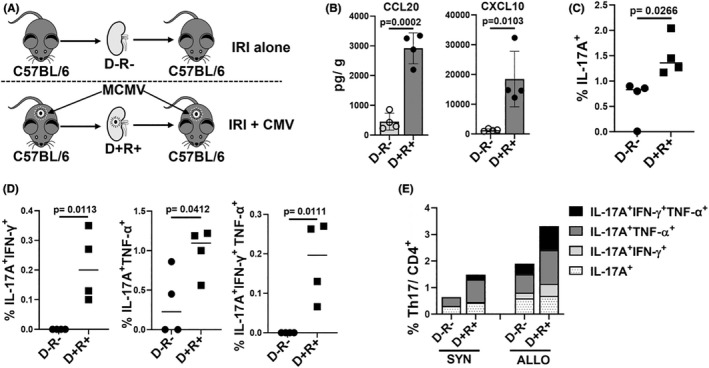
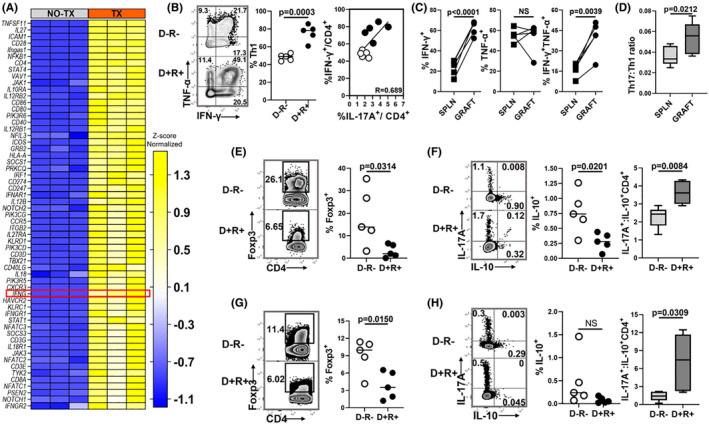
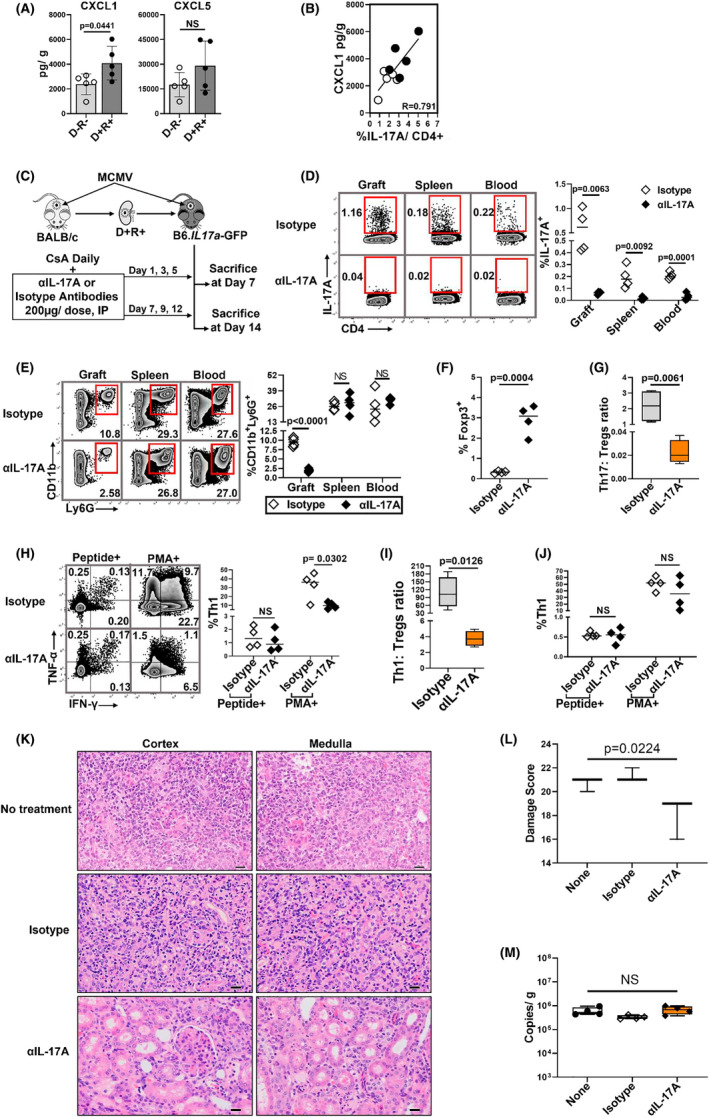
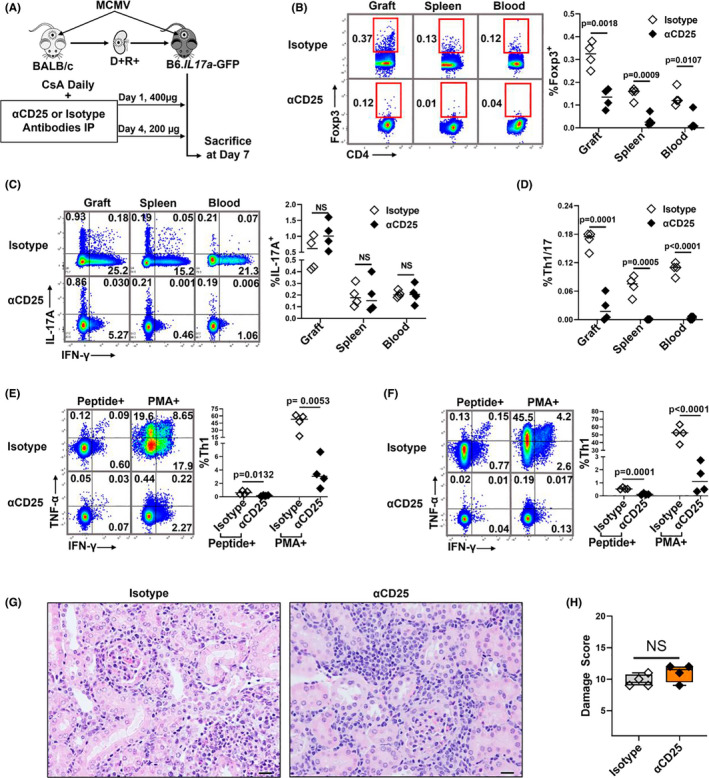
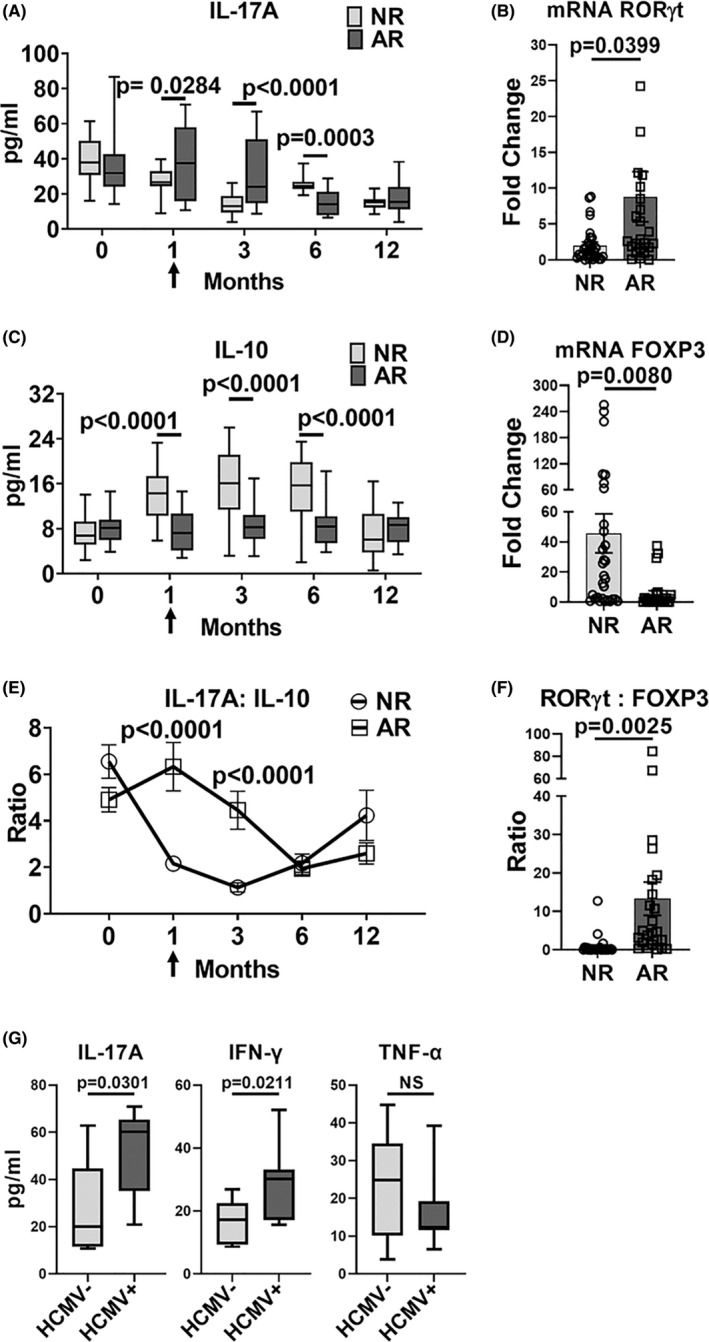
Human cytomegalovirus (HCMV) infection is associated with renal allograft failure. Allograft damage in animal models is accelerated by CMV-induced T helper 17 (Th17) cell infiltrates. However, the mechanisms whereby CMV promotes Th17 cell-mediated pathological organ inflammation are uncharacterized. Here we demonstrate that murine CMV (MCMV)-induced intragraft Th17 cells have a Th1/17 phenotype co-expressing IFN-γ and/or TNF-α, but only a minority of these cells are MCMV specific. Instead, MCMV promotes intragraft expression of CCL20 and CXCL10, which are associated with recruitment of CCR6+ CXCR3+ Th17 cells. MCMV also enhances Th17 cell infiltrates after ischemia-reperfusion injury, independent of allogeneic responses. Pharmacologic inhibition of the Th17 cell signature cytokine, IL-17A, ameliorates MCMV-associated allograft damage without increasing intragraft viral loads or reducing MCMV-specific Th1 cell infiltrates. Clinically, HCMV DNAemia is associated with higher serum IL-17A among renal transplant patients with acute rejection, linking HCMV reactivation with Th17 cell cytokine expression. In summary, CMV promotes allograft damage via cytokine-mediated Th1/17 cell recruitment, which may be pharmacologically targeted to mitigate graft injury while preserving antiviral T cell immunity.
Keywords: basic (laboratory) research/science; chemokines/chemokine receptors; clinical research/practice; infection and infectious agents-viral: cytomegalovirus (CMV); kidney transplantation/nephrology; rejection: acute, cytokines/cytokine receptors.
© 2022 The Authors. American Journal of Transplantation published by Wiley Periodicals LLC on behalf of The American Society of Transplantation and the American Society of Transplant Surgeons.
Figures