

3D cryo-EM imaging of bacterial flagella: Novel structural and mechanistic insights into cell motility
- PMID: 35671822
- PMCID: PMC9254593
- DOI: 10.1016/j.jbc.2022.102105
3D cryo-EM imaging of bacterial flagella: Novel structural and mechanistic insights into cell motility
Abstract
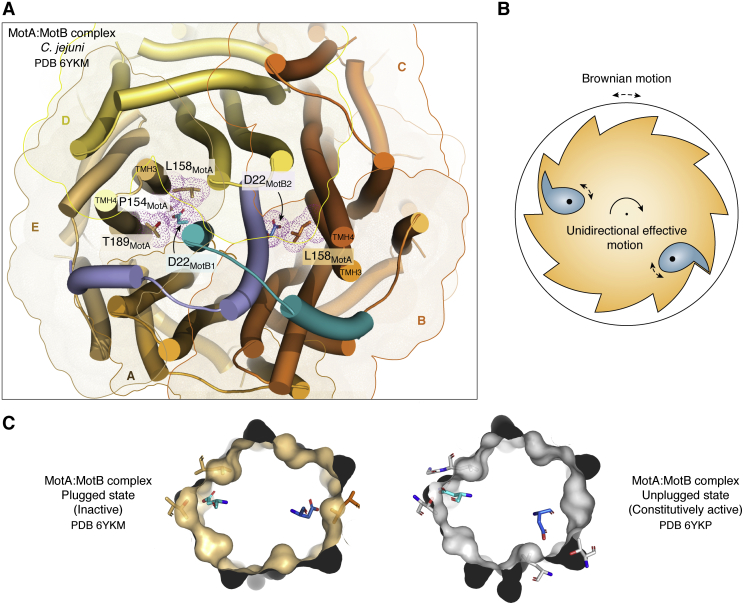
Bacterial flagella are nanomachines that enable cells to move at high speeds. Comprising 25 and more different types of proteins, the flagellum is a large supramolecular assembly organized into three widely conserved substructures: a basal body including the rotary motor, a connecting hook, and a long filament. The whole flagellum from Escherichia coli weighs ∼20 MDa, without considering its filament portion, which is by itself a ∼1.6 GDa structure arranged as a multimer of ∼30,000 flagellin protomers. Breakthroughs regarding flagellar structure and function have been achieved in the last few years, mainly because of the revolutionary improvements in 3D cryo-EM methods. This review discusses novel structures and mechanistic insights derived from such high-resolution studies, advancing our understanding of each one of the three major flagellar segments. The rotation mechanism of the motor has been unveiled with unprecedented detail, showing a two-cogwheel machine propelled by a Brownian ratchet device. In addition, by imaging the flagellin-like protomers that make up the hook in its native bent configuration, their unexpected conformational plasticity challenges the paradigm of a two-state conformational rearrangement mechanism for flagellin-fold proteins. Finally, imaging of the filaments of periplasmic flagella, which endow Spirochete bacteria with their singular motility style, uncovered a strikingly asymmetric protein sheath that coats the flagellin core, challenging the view of filaments as simple homopolymeric structures that work as freely whirling whips. Further research will shed more light on the functional details of this amazing nanomachine, but our current understanding has definitely come a long way.
Keywords: Spirochetes; allosteric regulation; cell motility; molecular motor; protein self-assembly; protein–protein interaction; proton motive force; proton transport; structural biology.
Copyright © 2022 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures





References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
