

Dynamic nucleosome landscape elicits a noncanonical GATA2 pioneer model
- PMID: 35672415
- PMCID: PMC9174260
- DOI: 10.1038/s41467-022-30960-x
Dynamic nucleosome landscape elicits a noncanonical GATA2 pioneer model
Abstract
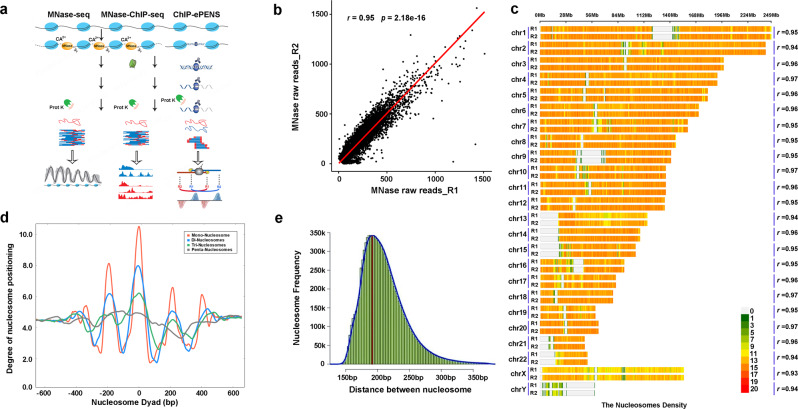
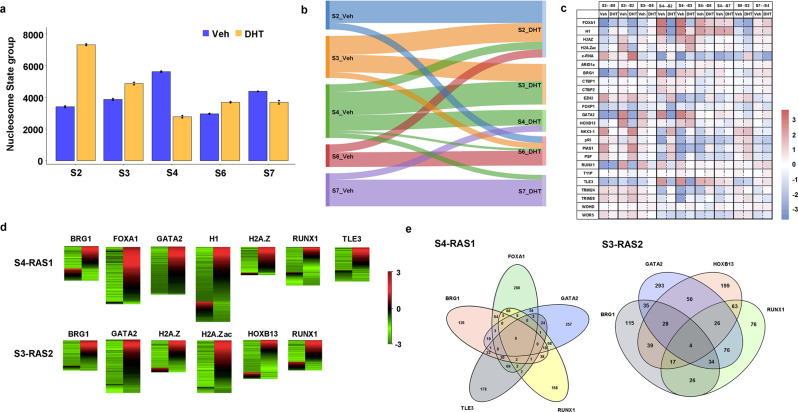
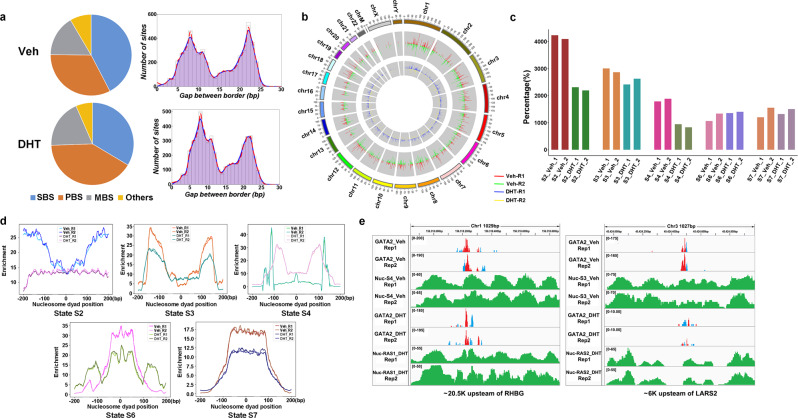
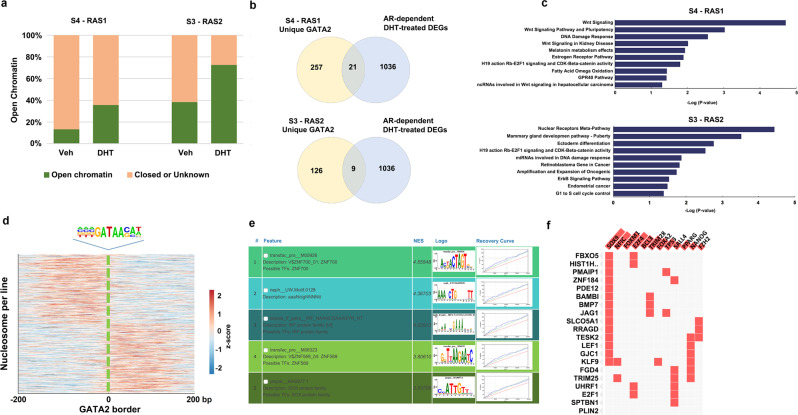
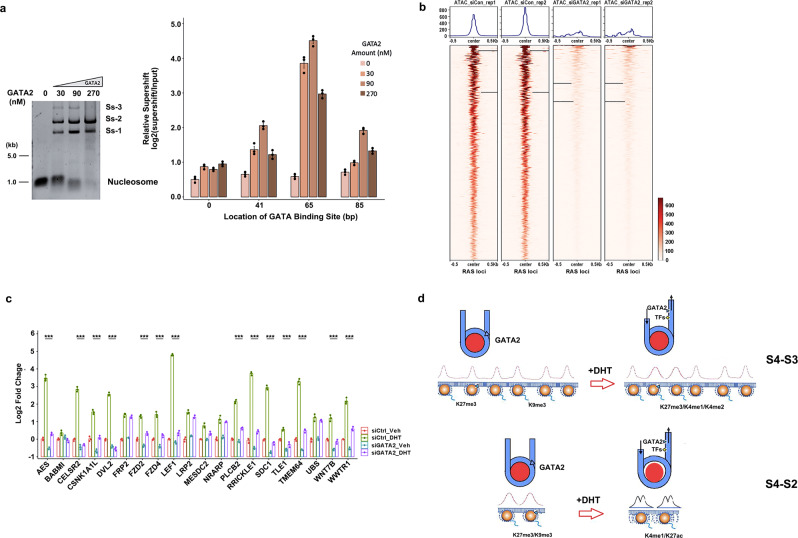
Knowledge gaps remain on how nucleosome organization and dynamic reorganization are governed by specific pioneer factors in a genome-wide manner. In this study, we generate over three billons of multi-omics sequencing data to exploit dynamic nucleosome landscape governed by pioneer factors (PFs), FOXA1 and GATA2. We quantitatively define nine functional nucleosome states each with specific characteristic nucleosome footprints in LNCaP prostate cancer cells. Interestingly, we observe dynamic switches among nucleosome states upon androgen stimulation, accompanied by distinct differential (gained or lost) binding of FOXA1, GATA2, H1 as well as many other coregulators. Intriguingly, we reveal a noncanonical pioneer model of GATA2 that it initially functions as a PF binding at the edge of a nucleosome in an inaccessible crowding array. Upon androgen stimulation, GATA2 re-configures an inaccessible to accessible nucleosome state and subsequently acts as a master transcription factor either directly or recruits signaling specific transcription factors to enhance WNT signaling in an androgen receptor (AR)-independent manner. Our data elicit a pioneer and master dual role of GATA2 in mediating nucleosome dynamics and enhancing downstream signaling pathways. Our work offers structural and mechanistic insight into the dynamics of pioneer factors governing nucleosome reorganization.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

Similar articles
-
Three-tiered role of the pioneer factor GATA2 in promoting androgen-dependent gene expression in prostate cancer.Nucleic Acids Res. 2014 Apr;42(6):3607-22. doi: 10.1093/nar/gkt1382. Epub 2014 Jan 13. Nucleic Acids Res. 2014. PMID: 24423874 Free PMC article.
-
Androgen receptor transcriptionally regulates semaphorin 3C in a GATA2-dependent manner.Oncotarget. 2017 Feb 7;8(6):9617-9633. doi: 10.18632/oncotarget.14168. Oncotarget. 2017. PMID: 28038451 Free PMC article.
-
Dynamic nucleosome-depleted regions at androgen receptor enhancers in the absence of ligand in prostate cancer cells.Mol Cell Biol. 2011 Dec;31(23):4648-62. doi: 10.1128/MCB.05934-11. Epub 2011 Oct 3. Mol Cell Biol. 2011. PMID: 21969603 Free PMC article.
-
Pioneer of prostate cancer: past, present and the future of FOXA1.Protein Cell. 2021 Jan;12(1):29-38. doi: 10.1007/s13238-020-00786-8. Epub 2020 Sep 18. Protein Cell. 2021. PMID: 32946061 Free PMC article. Review.
-
The role of GATA2 in lethal prostate cancer aggressiveness.Nat Rev Urol. 2017 Jan;14(1):38-48. doi: 10.1038/nrurol.2016.225. Epub 2016 Nov 22. Nat Rev Urol. 2017. PMID: 27872477 Free PMC article. Review.
Cited by
-
Induction of the zinc finger transcription factor GATA2 promotes kidney inflammation-related gene expression.J Biol Chem. 2025 Jul;301(7):110372. doi: 10.1016/j.jbc.2025.110372. Epub 2025 Jun 16. J Biol Chem. 2025. PMID: 40516868 Free PMC article.
-
Pioneer factors: roles and their regulation in development.Trends Genet. 2024 Feb;40(2):134-148. doi: 10.1016/j.tig.2023.10.007. Epub 2023 Nov 7. Trends Genet. 2024. PMID: 37940484 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
