

The roles of growth regulation and appendage patterning genes in the morphogenesis of treehopper pronota
- PMID: 35673859
- PMCID: PMC9174728
- DOI: 10.1098/rspb.2021.2682
The roles of growth regulation and appendage patterning genes in the morphogenesis of treehopper pronota
Abstract
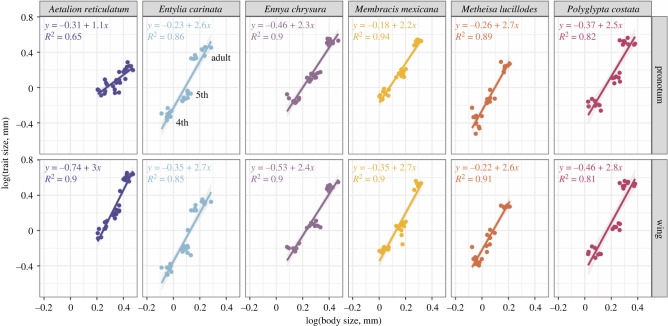
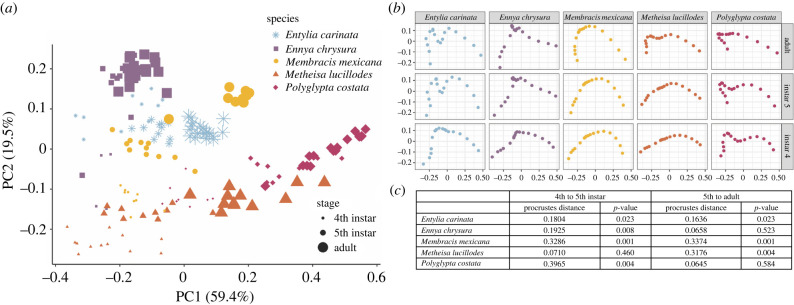
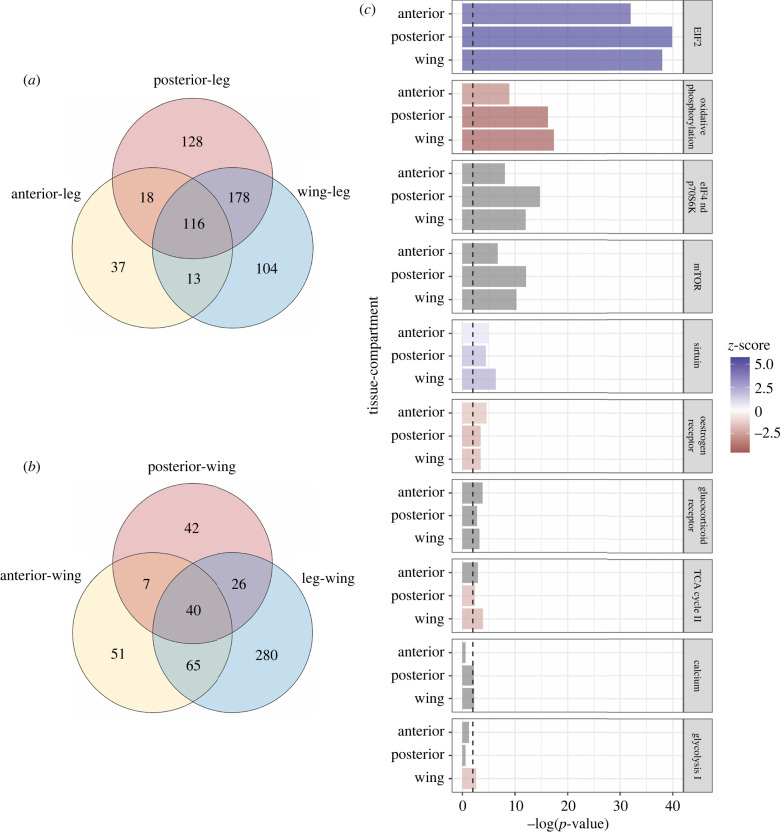
Treehoppers of the insect family Membracidae have evolved enlarged and elaborate pronotal structures, which is hypothesized to involve co-opted expression of genes that are shared with the wings. Here, we investigate the similarity between the pronotum and wings in relation to growth. Our study reveals that the ontogenetic allometry of the pronotum is similar to that of wings in Membracidae, but not the outgroup. Using transcriptomics, we identify genes related to translation and protein synthesis, which are mutually upregulated. These genes are implicated in the eIF2, eIF4/p70S6K and mTOR pathways, and have known roles in regulating cell growth and proliferation. We find that species-specific differential growth patterning of the pronotum begins as early as the third instar, which suggests that expression of appendage patterning genes occurs long before the metamorphic molt. We propose that a network related to growth and size determination is the more likely mechanism shared with wings. However, regulators upstream of the shared genes in pronotum and wings need to be elucidated to substantiate whether co-option has occurred. Finally, we believe it will be helpful to distinguish the mechanisms leading to pronotal size from those regulating pronotal shape as we make sense of this spectacular evolutionary innovation.
Keywords: Entylia carinata; growth; membracidae; morphogenesis; pronotum; treehopper.
Conflict of interest statement
There are no competing interests.
Figures

Similar articles
-
Ontogenetic trajectories and early shape differentiation of treehopper pronota (Hemiptera: Membracidae).Evol Dev. 2023 May;25(3):240-252. doi: 10.1111/ede.12431. Epub 2023 Apr 10. Evol Dev. 2023. PMID: 37035938
-
Co-option of wing-patterning genes underlies the evolution of the treehopper helmet.Nat Ecol Evol. 2020 Feb;4(2):250-260. doi: 10.1038/s41559-019-1054-4. Epub 2019 Dec 9. Nat Ecol Evol. 2020. PMID: 31819237
-
Structure and development of the complex helmet of treehoppers (Insecta: Hemiptera: Membracidae).Zoological Lett. 2020 Feb 24;6:3. doi: 10.1186/s40851-020-00155-7. eCollection 2020. Zoological Lett. 2020. PMID: 32123574 Free PMC article.
-
Morphogenesis in bat wings: linking development, evolution and ecology.Cells Tissues Organs. 2008;187(1):13-23. doi: 10.1159/000109960. Cells Tissues Organs. 2008. PMID: 18163246 Review.
-
Temperature coupling of mate attraction signals and female mate preferences in four populations of Enchenopa treehopper (Hemiptera: Membracidae).J Evol Biol. 2019 Oct;32(10):1046-1056. doi: 10.1111/jeb.13506. Epub 2019 Jul 30. J Evol Biol. 2019. PMID: 31278803 Review.
Cited by
-
Electroreception in treehoppers: How extreme morphologies can increase electrical sensitivity.Proc Natl Acad Sci U S A. 2025 Jul 29;122(30):e2505253122. doi: 10.1073/pnas.2505253122. Epub 2025 Jul 21. Proc Natl Acad Sci U S A. 2025. PMID: 40690666 Free PMC article.
References
-
- Wood TK. 1993. Diversity in the New World Membracidae. Annu. Rev. Entomol. 38, 409-433. (10.1146/annurev.en.38.010193.002205) - DOI
-
- Godoy C, Miranda X, Nishida K.. 2006. Treehoppers of Tropical America/Membrácidos de la América Tropical, 1st edn. Santo Domingo, Heredia, Costa Rica. Instituto; Nacional de Biodiversidad. See https://www.nhbs.com/treehoppers-of-tropical-america-membracidos-de-la-a....
-
- Evangelista O, Sakakibara AM, Cryan JR, Urban JM. 2017. A phylogeny of the treehopper subfamily Heteronotinae reveals convergent pronotal traits (Hemiptera: Auchenorrhyncha: Membracidae). Syst. Entomol. 42, 410-428. (10.1111/syen.12221) - DOI
Publication types
MeSH terms
Associated data
LinkOut - more resources
Full Text Sources
Miscellaneous
