

Dynamic interplay between the periplasmic chaperone SurA and the BAM complex in outer membrane protein folding
- PMID: 35676411
- PMCID: PMC9177699
- DOI: 10.1038/s42003-022-03502-w
Dynamic interplay between the periplasmic chaperone SurA and the BAM complex in outer membrane protein folding
Abstract
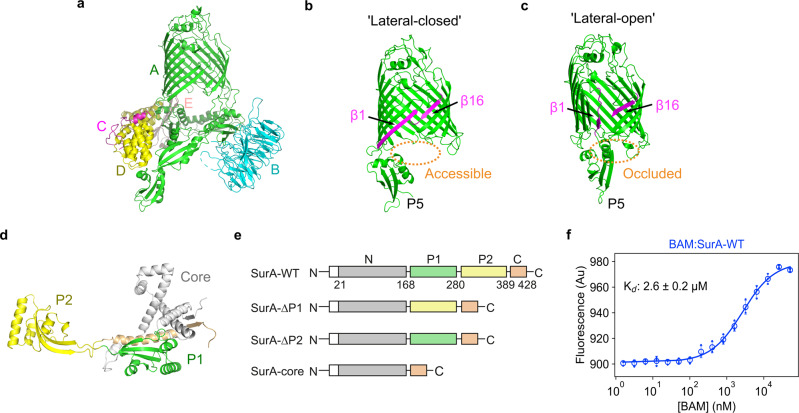
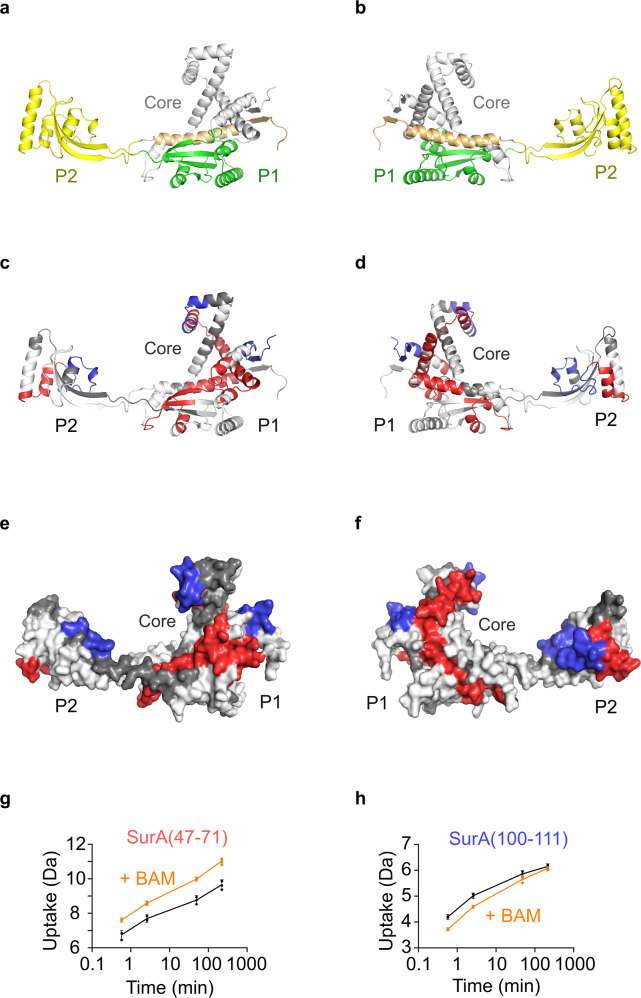
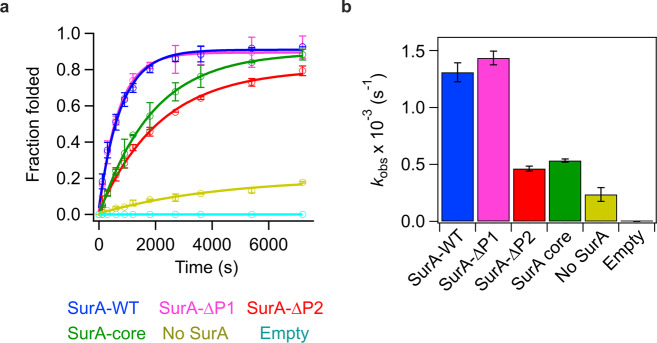
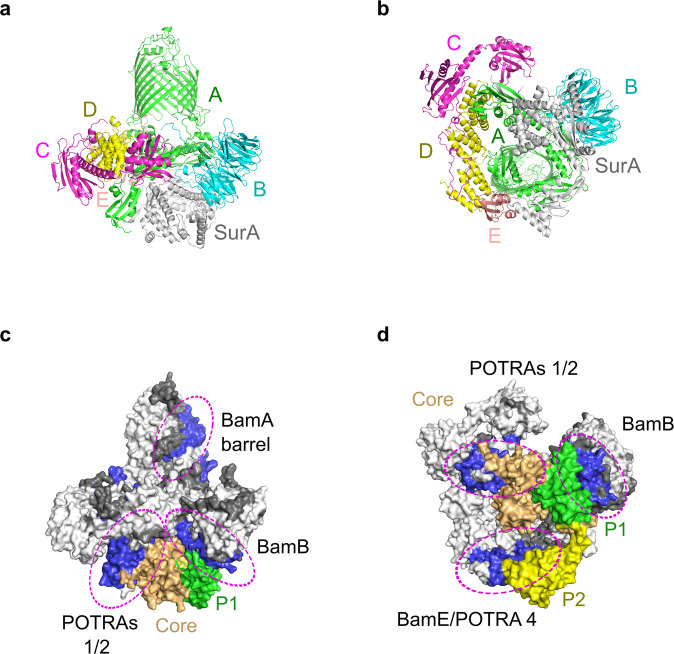
Correct folding of outer membrane proteins (OMPs) into the outer membrane of Gram-negative bacteria depends on delivery of unfolded OMPs to the β-barrel assembly machinery (BAM). How unfolded substrates are presented to BAM remains elusive, but the major OMP chaperone SurA is proposed to play a key role. Here, we have used hydrogen deuterium exchange mass spectrometry (HDX-MS), crosslinking, in vitro folding and binding assays and computational modelling to show that the core domain of SurA and one of its two PPIase domains are key to the SurA-BAM interaction and are required for maximal catalysis of OMP folding. We reveal that binding causes changes in BAM and SurA conformation and/or dynamics distal to the sites of binding, including at the BamA β1-β16 seam. We propose a model for OMP biogenesis in which SurA plays a crucial role in OMP delivery and primes BAM to accept substrates for folding.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


Similar articles
-
Outer membrane protein assembly mediated by BAM-SurA complexes.Nat Commun. 2024 Sep 1;15(1):7612. doi: 10.1038/s41467-024-51358-x. Nat Commun. 2024. PMID: 39218969 Free PMC article.
-
Dual client binding sites in the ATP-independent chaperone SurA.Nat Commun. 2024 Sep 14;15(1):8071. doi: 10.1038/s41467-024-52021-1. Nat Commun. 2024. PMID: 39277579 Free PMC article.
-
Effects of Periplasmic Chaperones and Membrane Thickness on BamA-Catalyzed Outer-Membrane Protein Folding.J Mol Biol. 2017 Nov 24;429(23):3776-3792. doi: 10.1016/j.jmb.2017.09.008. Epub 2017 Sep 15. J Mol Biol. 2017. PMID: 28919234 Free PMC article.
-
From Chaperones to the Membrane with a BAM!Trends Biochem Sci. 2016 Oct;41(10):872-882. doi: 10.1016/j.tibs.2016.06.005. Epub 2016 Jul 19. Trends Biochem Sci. 2016. PMID: 27450425 Free PMC article. Review.
-
The role of SurA factor in outer membrane protein transport and virulence.Int J Med Microbiol. 2010 Nov;300(7):421-8. doi: 10.1016/j.ijmm.2010.04.012. Epub 2010 May 6. Int J Med Microbiol. 2010. PMID: 20447864 Review.
Cited by
-
FkpA enhances membrane protein folding using an extensive interaction surface.Protein Sci. 2023 Apr;32(4):e4592. doi: 10.1002/pro.4592. Protein Sci. 2023. PMID: 36775935 Free PMC article.
-
Lateral gating mechanism and plasticity of the β-barrel assembly machinery complex in micelles and Escherichia coli.PNAS Nexus. 2024 Jan 17;3(2):pgae019. doi: 10.1093/pnasnexus/pgae019. eCollection 2024 Feb. PNAS Nexus. 2024. PMID: 38312222 Free PMC article.
-
SurA-like and Skp-like Proteins as Important Virulence Determinants of the Gram Negative Bacterial Pathogens.Int J Mol Sci. 2022 Dec 24;24(1):295. doi: 10.3390/ijms24010295. Int J Mol Sci. 2022. PMID: 36613738 Free PMC article. Review.
-
Analysis of Transmembrane β-Barrel Proteins by Native and Semi-native Polyacrylamide Gel Electrophoresis.Methods Mol Biol. 2024;2778:133-145. doi: 10.1007/978-1-0716-3734-0_9. Methods Mol Biol. 2024. PMID: 38478276
-
The Name Is Barrel, β-Barrel.Methods Mol Biol. 2024;2778:1-30. doi: 10.1007/978-1-0716-3734-0_1. Methods Mol Biol. 2024. PMID: 38478268
References
Publication types
MeSH terms
Substances
Grants and funding
- BB/P000037/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- 220628/Z/20/Z/WT_/Wellcome Trust/United Kingdom
- BB/N007603/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/M012573/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- MR/T011149/1/MRC_/Medical Research Council/United Kingdom
- 223268/Z/21/Z/WT_/Wellcome Trust/United Kingdom
- 220628/WT_/Wellcome Trust/United Kingdom
- 105615/Z/14/Z/WT_/Wellcome Trust/United Kingdom
- NIHR200633/DH_/Department of Health/United Kingdom
- BB/T000635/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- 222373/Z/21/Z/WT_/Wellcome Trust/United Kingdom
- 208385/Z/17/Z/WT_/Wellcome Trust/United Kingdom
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
