

Genome evolution and diversity of wild and cultivated potatoes
- PMID: 35676481
- PMCID: PMC9200641
- DOI: 10.1038/s41586-022-04822-x
Genome evolution and diversity of wild and cultivated potatoes
Abstract
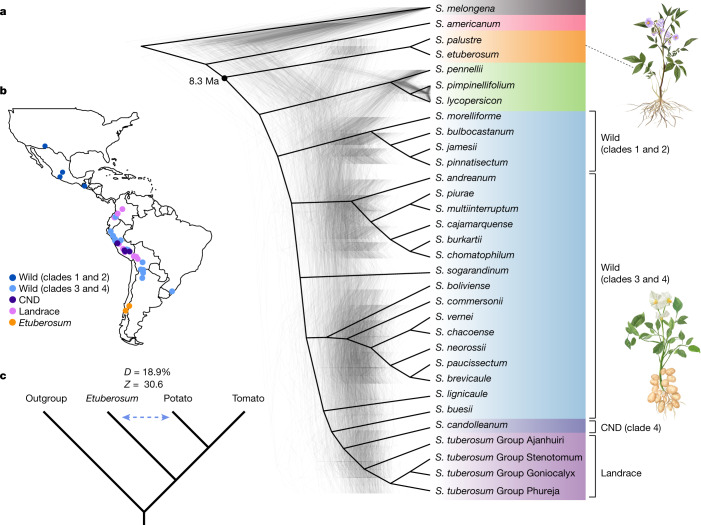
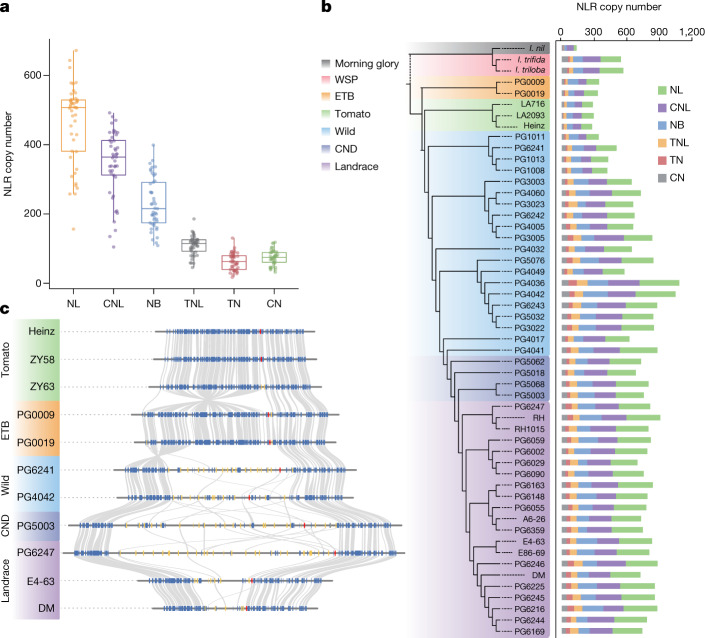
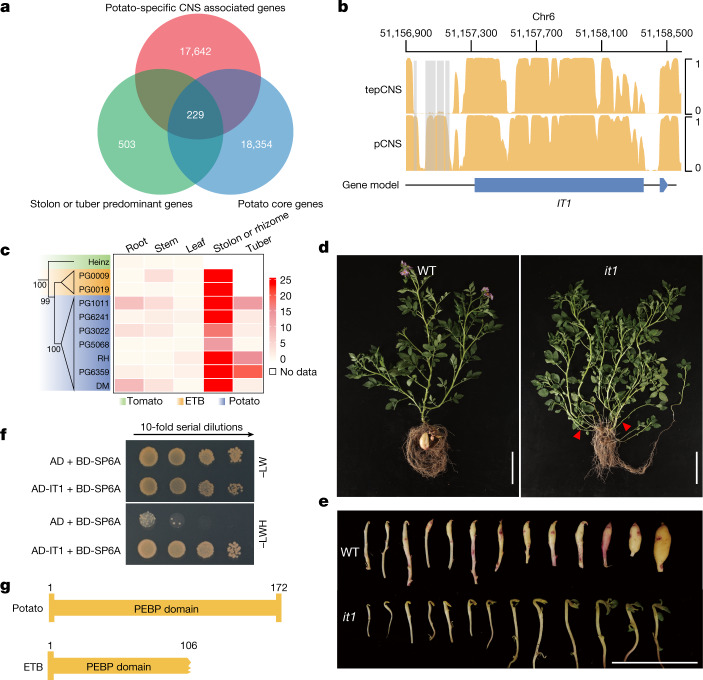
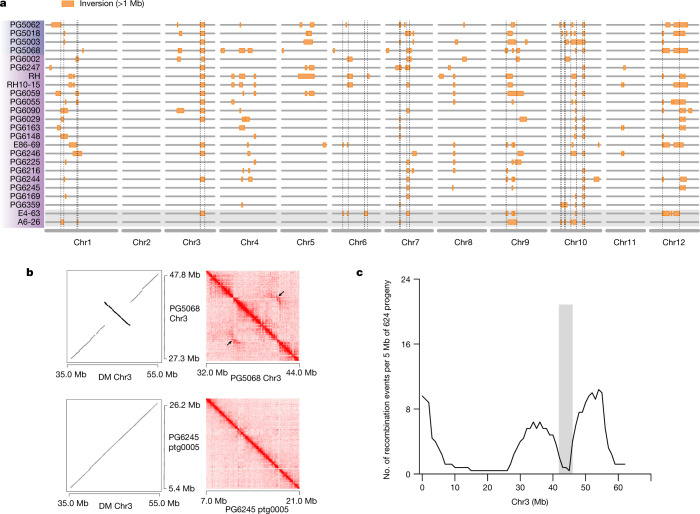
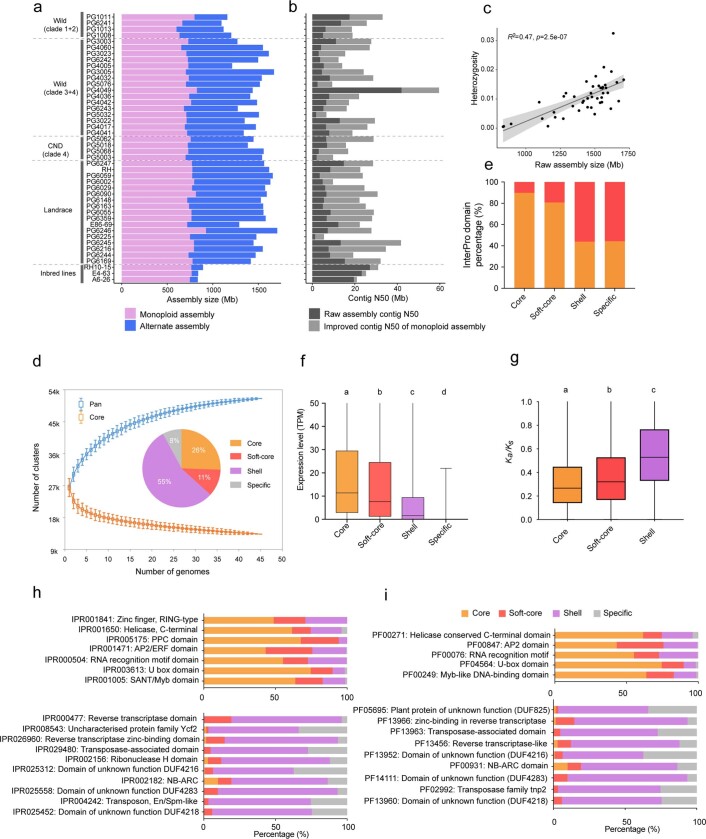
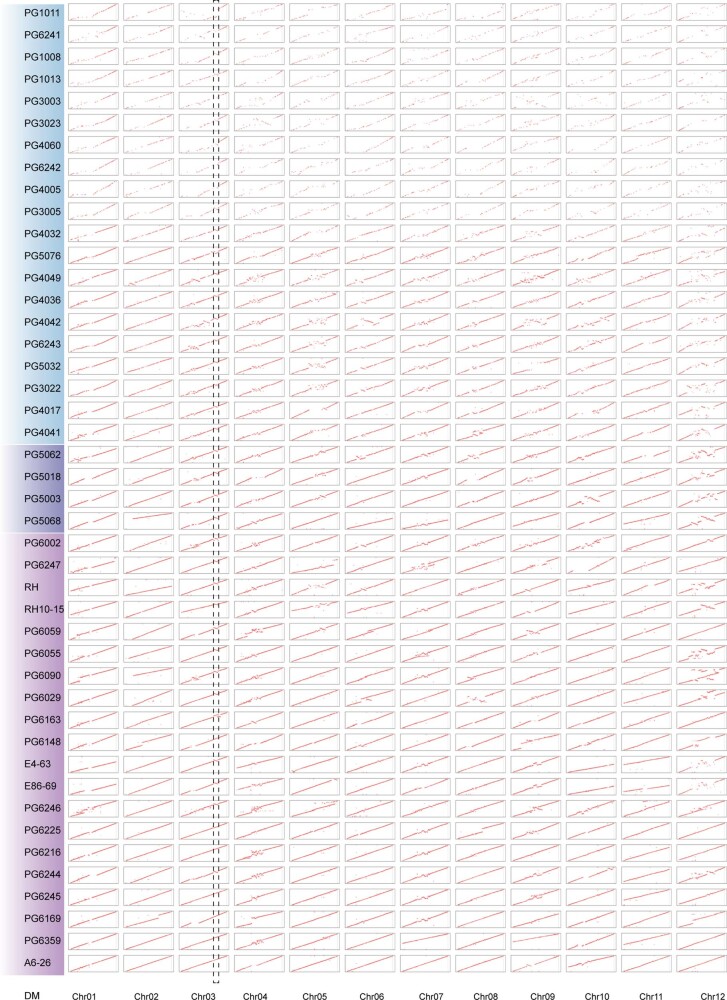
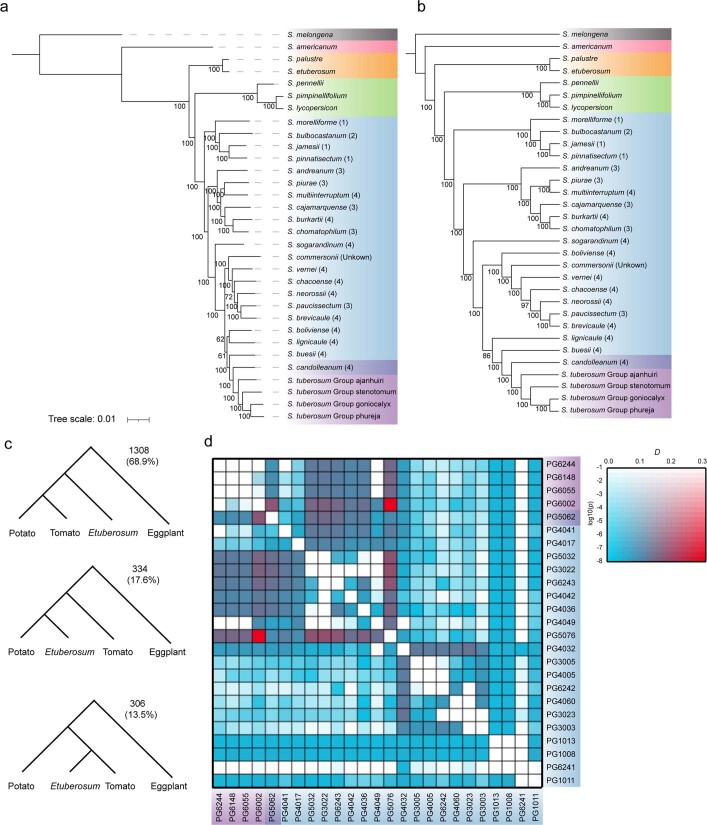
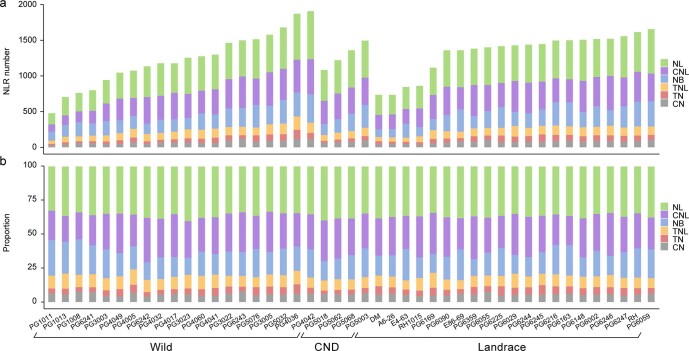
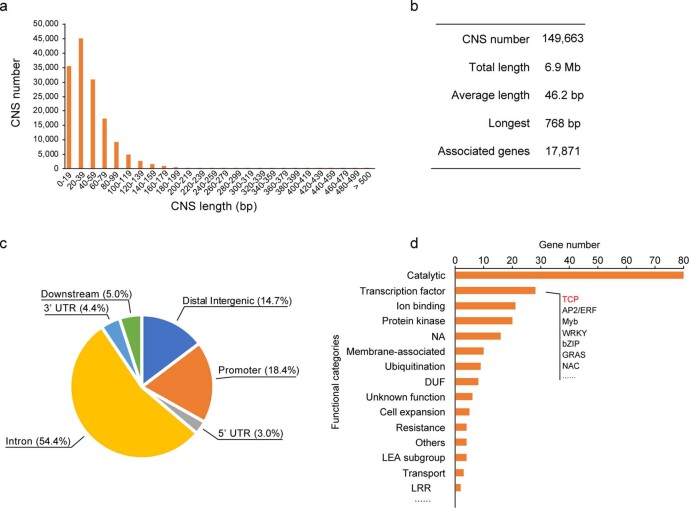
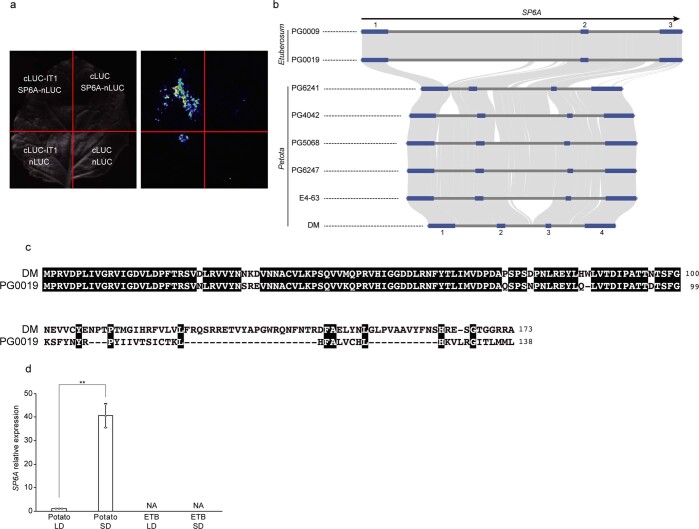
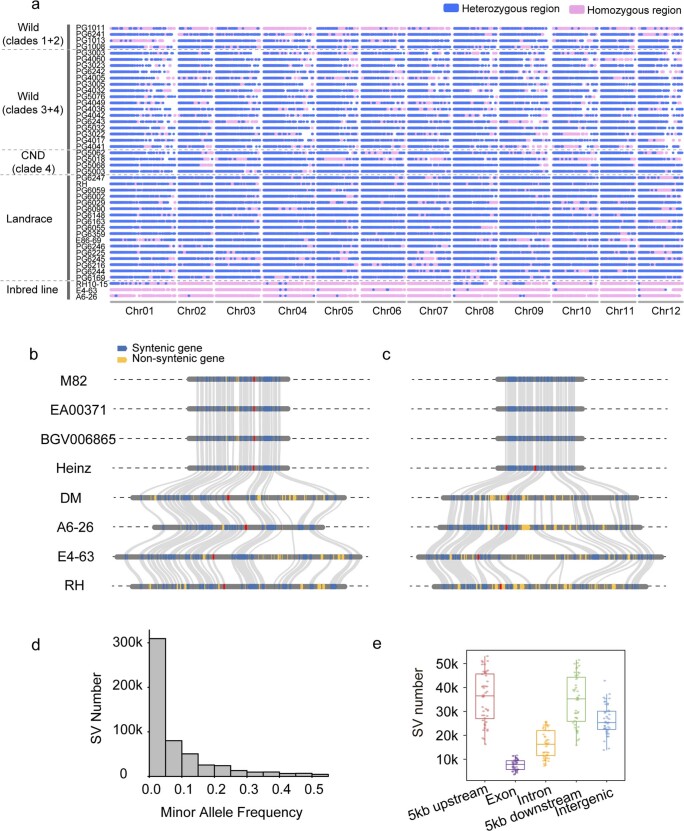
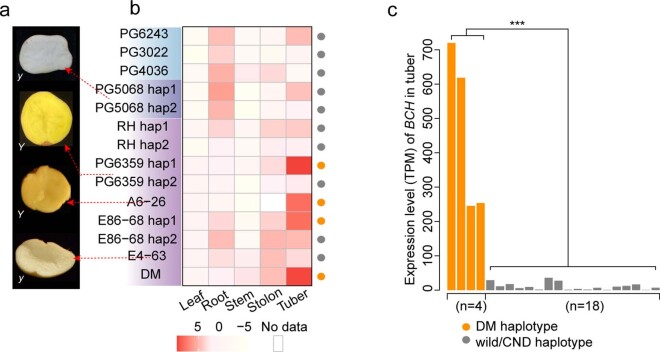
Potato (Solanum tuberosum L.) is the world's most important non-cereal food crop, and the vast majority of commercially grown cultivars are highly heterozygous tetraploids. Advances in diploid hybrid breeding based on true seeds have the potential to revolutionize future potato breeding and production1-4. So far, relatively few studies have examined the genome evolution and diversity of wild and cultivated landrace potatoes, which limits the application of their diversity in potato breeding. Here we assemble 44 high-quality diploid potato genomes from 24 wild and 20 cultivated accessions that are representative of Solanum section Petota, the tuber-bearing clade, as well as 2 genomes from the neighbouring section, Etuberosum. Extensive discordance of phylogenomic relationships suggests the complexity of potato evolution. We find that the potato genome substantially expanded its repertoire of disease-resistance genes when compared with closely related seed-propagated solanaceous crops, indicative of the effect of tuber-based propagation strategies on the evolution of the potato genome. We discover a transcription factor that determines tuber identity and interacts with the mobile tuberization inductive signal SP6A. We also identify 561,433 high-confidence structural variants and construct a map of large inversions, which provides insights for improving inbred lines and precluding potential linkage drag, as exemplified by a 5.8-Mb inversion that is associated with carotenoid content in tubers. This study will accelerate hybrid potato breeding and enrich our understanding of the evolution and biology of potato as a global staple food crop.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

Comment in
-
Potato genomes pave the way to crop improvement.Nature. 2022 Jun;606(7914):472-473. doi: 10.1038/d41586-022-01419-2. Nature. 2022. PMID: 35676347 No abstract available.
-
Insights of section-wide pan-genome into hybrid potato breeding.Sci China Life Sci. 2022 Oct;65(10):2125-2127. doi: 10.1007/s11427-022-2144-9. Epub 2022 Jun 21. Sci China Life Sci. 2022. PMID: 35752693 No abstract available.
References
-
- Lindhout P, et al. Towards F1 hybrid seed potato breeding. Potato Res. 2011;54:301–312. doi: 10.1007/s11540-011-9196-z. - DOI
-
- Li Y, Li G, Li C, Qu D, Huang S. Prospects of diploid hybrid breeding in potato. Chinese Potato J. 2013;27:96–99.
-
- Spooner DM, Ghislain M, Simon R, Jansky SH, Gavrilenko T. Systematics, diversity, genetics, and evolution of wild and cultivated potatoes. Bot. Rev. 2014;80:283–383. doi: 10.1007/s12229-014-9146-y. - DOI
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
