

The influence of temporal unpredictability on the electrophysiological mechanisms of neural entrainment
- PMID: 35678104
- PMCID: PMC9787398
- DOI: 10.1111/psyp.14108
The influence of temporal unpredictability on the electrophysiological mechanisms of neural entrainment
Abstract
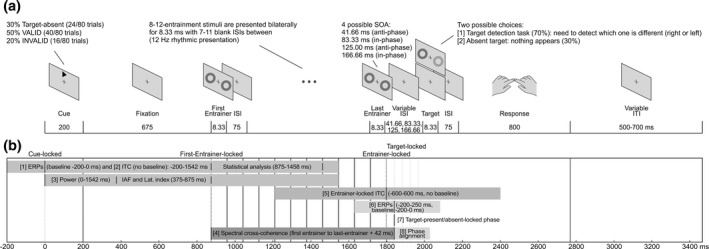
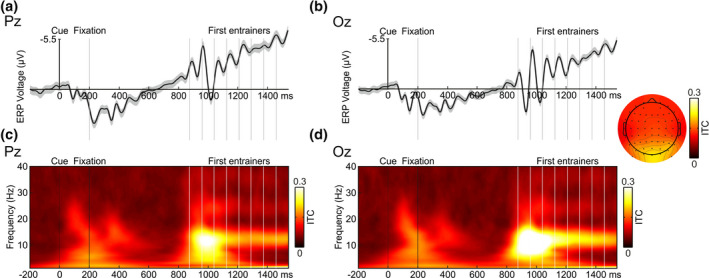
Neural entrainment, or the synchronization of endogenous oscillations to exogenous rhythmic events, has been postulated as a powerful mechanism underlying stimulus prediction. Nevertheless, studies that have explored the benefits of neural entrainment on attention, perception, and other cognitive functions have received criticism, which could compromise their theoretical and clinical value. Therefore, the aim of the present study was [1] to confirm the presence of entrainment using a set of pre-established criteria and [2] to establish whether the reported behavioral benefits of entrainment remain when temporal predictability related to target appearance is reduced. To address these points, we adapted a previous neural entrainment paradigm to include: a variable entrainer length and increased target-absent trials, and instructing participants to respond only if they had detected a target, to avoid guessing. Thirty-six right-handed women took part in this study. Our results indicated a significant alignment of neural activity to the external periodicity as well as a persistence of phase alignment beyond the offset of the driving signal. This would appear to indicate that neural entrainment triggers preexisting endogenous oscillations, which cannot simply be explained as a succession of event-related potentials associated with the stimuli, expectation and/or motor response. However, we found no behavioral benefit for targets in-phase with entrainers, which would suggest that the effect of neural entrainment on overt behavior may be more limited than expected. These results help to clarify the mechanistic processes underlying neural entrainment and provide new insights on its applications.
Keywords: EEG; alpha rhythm; endogenous oscillations; entrainment; phase synchronization; temporal unpredictability.
© 2022 The Authors. Psychophysiology published by Wiley Periodicals LLC on behalf of Society for Psychophysiological Research.
Conflict of interest statement
The project leading to these results has received funding from “la Caixa” Foundation under the project code LCF/PR/PR16/51110005 and has been co‐funded with 50% by the European Regional Development Fund under the framework of the ERFD Operative Programme for Catalunya 2014–2020, with a grant of 1.527.637,88€. AVB is supported by the Spanish MICINN Juan de la Cierva postdoctoral grant (FJC2018‐036804‐I), and AMM by a predoctoral grant from the “Fundació Institut de Recerca Hospital Universitari Vall d'Hebron” (VHIR/BEQUESPREDOC/2020/MARTI). AVB and AMM have received a postdoctoral and a predoctoral contract respectively, from the “La Caixa” Foundation. SS‐F is supported by grants from the Ministerio de Ciencia e Innovación (PID2019‐108531GB‐I00 AEI/FEDER) and AGAUR Generalitat de Catalunya (2017 SGR 1545). PPR has received honoraria as a consultant and speaker for Allergan, Almirall, Biohaven, Chiesi, Eli Lilly, Medscape, Neurodiem, Novartis, and Teva. Her research group has received research grants from Allergan, AGAUR, la Caixa foundation, Migraine Research Foundation, Instituto Investigación Carlos III, MICINN, PERIS; and has received funding for clinical trials from Alder, Electrocore, Eli Lilly, Novartis, and Teva. She is a trustee member of the board of the International Headache Society and a member of the Council of the European Headache Federation. She is on the editorial board of Revista de Neurologia. She is an editor for Frontiers of Neurology and the Journal of Headache and Pain. She is a member of the Clinical Trials Guidelines Committee of the International Headache Society. She has edited the Guidelines for the Diagnosis and Treatment of Headache of the Spanish Neurological Society. She is the founder of
Figures




References
-
- Beck, A. , Steer, R. , & Brown, G. (Eds.). (1996). Manual for the Beck Depression Inventory‐II. Psychological Corporation.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
