

Robust variation in infant gut microbiome assembly across a spectrum of lifestyles
- PMID: 35679413
- PMCID: PMC9894631
- DOI: 10.1126/science.abj2972
Robust variation in infant gut microbiome assembly across a spectrum of lifestyles
Abstract
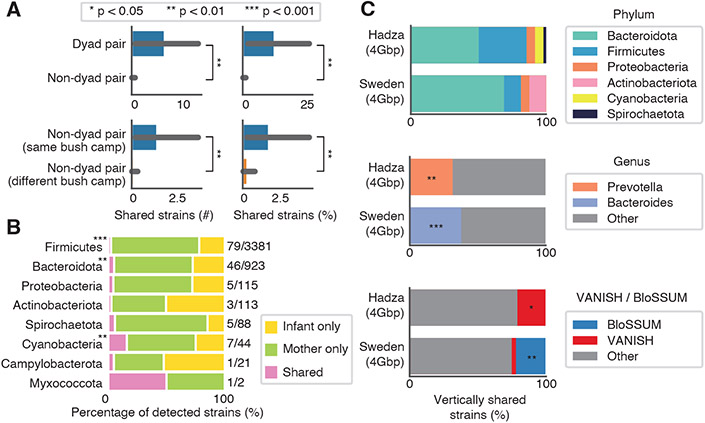
Infant microbiome assembly has been intensely studied in infants from industrialized nations, but little is known about this process in nonindustrialized populations. We deeply sequenced infant stool samples from the Hadza hunter-gatherers of Tanzania and analyzed them in a global meta-analysis. Infant microbiomes develop along lifestyle-associated trajectories, with more than 20% of genomes detected in the Hadza infant gut representing novel species. Industrialized infants-even those who are breastfed-have microbiomes characterized by a paucity of Bifidobacterium infantis and gene cassettes involved in human milk utilization. Strains within lifestyle-associated taxonomic groups are shared between mother-infant dyads, consistent with early life inheritance of lifestyle-shaped microbiomes. The population-specific differences in infant microbiome composition and function underscore the importance of studying microbiomes from people outside of wealthy, industrialized nations.
Figures


References
-
- Yatsunenko T, Rey FE, Manary MJ, Trehan I, Dominguez-Bello MG, Contreras M, Magris M, Hidalgo G, Baldassano RN, Anokhin AP, Heath AC, Warner B, Reeder J, Kuczynski J, Caporaso JG, Lozupone CA, Lauber C, Clemente JC, Knights D, Knight R, Gordon JI, Human gut microbiome viewed across age and geography. Nature. 486, 222–227 (2012). - PMC - PubMed
-
- Stewart CJ, Ajami NJ, O’Brien JL, Hutchinson DS, Smith DP, Wong MC, Ross MC, Lloyd RE, Doddapaneni H, Metcalf GA, Muzny D, Gibbs RA, Vatanen T, Huttenhower C, Xavier RJ, Rewers M, Hagopian W, Toppari J, Ziegler A-G, She J-X, Akolkar B, Lernmark A, Hyoty H, Vehik K, Krischer JP, Petrosino JF, Temporal development of the gut microbiome in early childhood from the TEDDY study. Nature. 562, 583–588 (2018). - PMC - PubMed
-
- de Goffau MC, Jallow AT, Sanyang C, Prentice AM, Meagher N, Price DJ, Revill PA, Parkhill J, Pereira DIA, Wagner J, Gut microbiomes from Gambian infants reveal the development of a non-industrialized Prevotella-based trophic network. Nat Microbiol (2021), doi: 10.1038/s41564-021-01023-6. - DOI - PMC - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
