

Direct epitranscriptomic regulation of mammalian translation initiation through N4-acetylcytidine
- PMID: 35679869
- PMCID: PMC9361928
- DOI: 10.1016/j.molcel.2022.05.016
Direct epitranscriptomic regulation of mammalian translation initiation through N4-acetylcytidine
Erratum in
-
Direct epitranscriptomic regulation of mammalian translation initiation through N4-acetylcytidine.Mol Cell. 2022 Aug 4;82(15):2912. doi: 10.1016/j.molcel.2022.06.022. Mol Cell. 2022. PMID: 35931039 Free PMC article. No abstract available.
Abstract
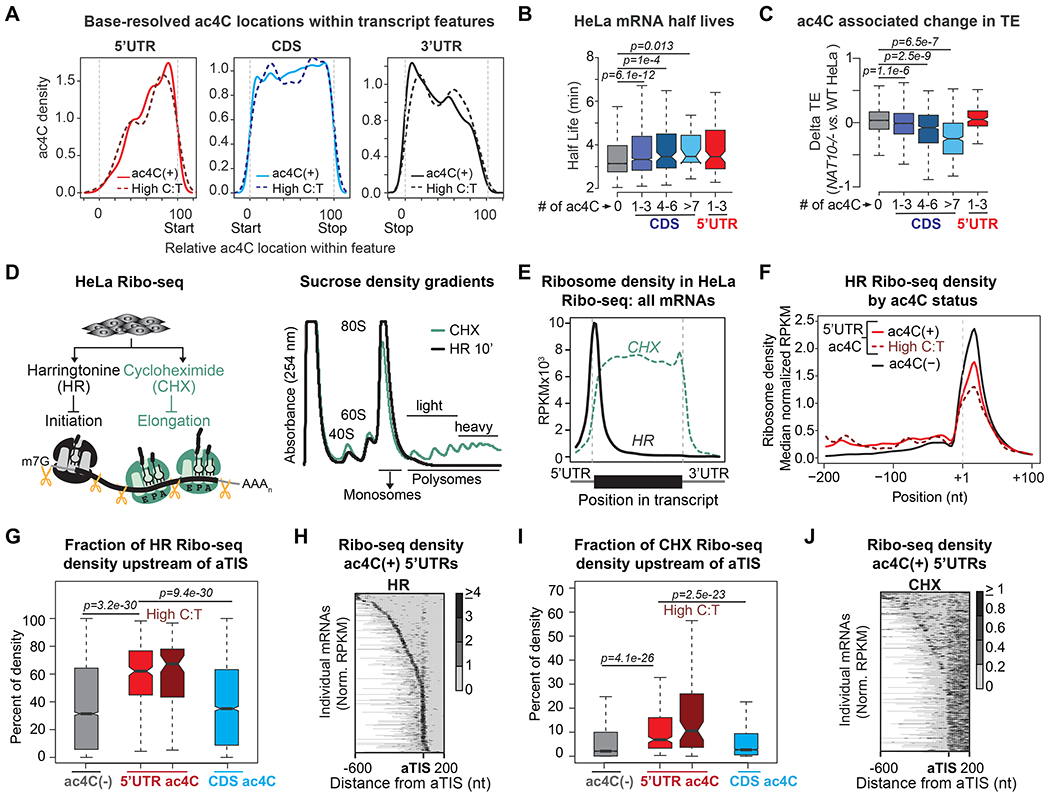
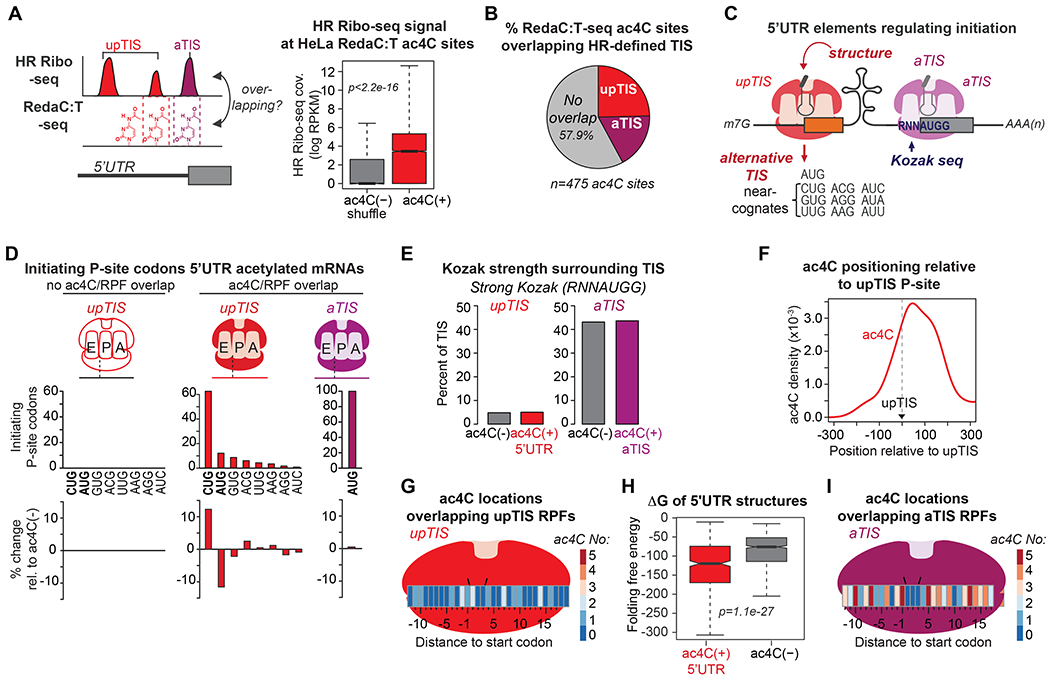
mRNA function is influenced by modifications that modulate canonical nucleobase behavior. We show that a single modification mediates distinct impacts on mRNA translation in a position-dependent manner. Although cytidine acetylation (ac4C) within protein-coding sequences stimulates translation, ac4C within 5' UTRs impacts protein synthesis at the level of initiation. 5' UTR acetylation promotes initiation at upstream sequences, competitively inhibiting annotated start codons. Acetylation further directly impedes initiation at optimal AUG contexts: ac4C within AUG-flanking Kozak sequences reduced initiation in base-resolved transcriptome-wide HeLa results and in vitro utilizing substrates with site-specific ac4C incorporation. Cryo-EM of mammalian 80S initiation complexes revealed that ac4C in the -1 position adjacent to an AUG start codon disrupts an interaction between C and hypermodified t6A at nucleotide 37 of the initiator tRNA. These findings demonstrate the impact of RNA modifications on nucleobase function at a molecular level and introduce mRNA acetylation as a factor regulating translation in a location-specific manner.
Keywords: 80S; Kozak; NAT10; ac4C; acetylcytidine; cryo-EM; epitranscriptome; initiation; t6A; translation.
Published by Elsevier Inc.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures





References
-
- Afonine PV, Grosse-Kunstleve RW, Echols N, Headd JJ, Moriarty NW, Mustyakimov M, Terwilliger TC, Urzhumtsev A, Zwart PH, and Adams PD (2012). Towards automated crystallographic structure refinement with phenix.refine. Acta Crystallogr D Biol Crystallogr 68, 352–367. 10.1107/S0907444912001308. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
