

CkREV Enhances the Drought Resistance of Caragana korshinskii through Regulating the Expression of Auxin Synthetase Gene CkYUC5
- PMID: 35682582
- PMCID: PMC9180416
- DOI: 10.3390/ijms23115902
CkREV Enhances the Drought Resistance of Caragana korshinskii through Regulating the Expression of Auxin Synthetase Gene CkYUC5
Abstract
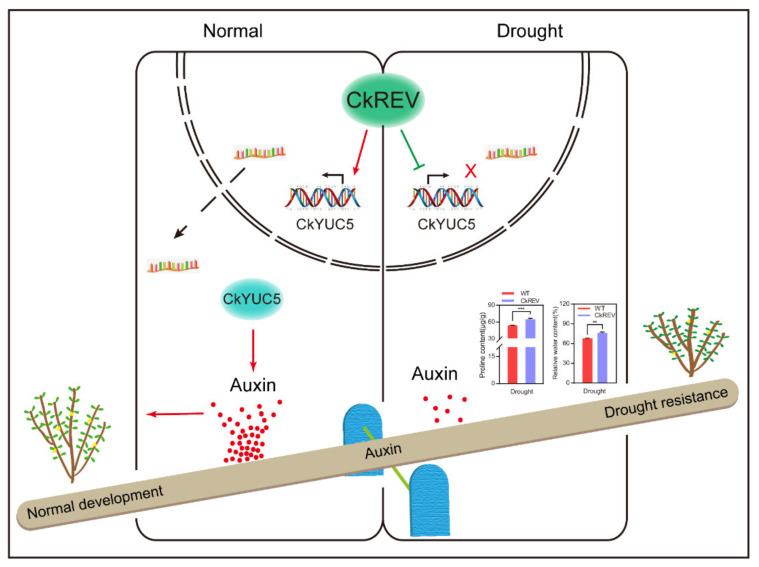
As a common abiotic stress, drought severely impairs the growth, development, and even survival of plants. Here we report a transcription factor, Caragana korshinskii REVOLUTA(CkREV), which can bidirectionally regulate the expression of the critical enzyme gene CkYUC5 in auxin synthesis according to external environment changes, so as to control the biosynthesis of auxin and further enhance the drought resistance of plants. Quantitative analysis reveals that the expression level of both CkYUC5 and AtYUC5 is down-regulated after C. korshinskii and Arabidopsis thaliana are exposed to drought. Functional verification of CkREV reveals that CkREV up-regulates the expression of AtYUC5 in transgenic A. thaliana under common conditions, while down-regulating it under drought conditions. Meanwhile, the expression of CkYUC5 is also down-regulated in C. korshinskii leaves instantaneously overexpressing CkREV. We apply a dual-luciferase reporter system to discover that CkREV can bind to the promoter of CkYUC5 to regulate its expression, which is further proved by EMSA and Y1H esxperiments. Functional verification of CkREV in C. korshinskii and transgenic A. thaliana shows that CkREV can regulate the expression of CkYUC5 and AtYUC5 in a contrary way, maintaining the equilibrium of plants between growth and drought resisting. CkREV can positively regulate the expression of CkYUC5 to promote auxin synthesis in favor of growth under normal development. However, CkREV can also respond to external signals and negatively regulate the expression of CkYUC5, which inhibits auxin synthesis in order to reduce growth rate, lower water demands, and eventually improve the drought resistance of plants.
Keywords: Caragana korshinskii; HD-ZIP III; auxin synthesis; drought; stress resistance.
Conflict of interest statement
The authors declare no conflict of interest.
Figures







References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
