

Integrate Small RNA and Degradome Sequencing to Reveal Drought Memory Response in Wheat (Triticum aestivum L.)
- PMID: 35682597
- PMCID: PMC9180835
- DOI: 10.3390/ijms23115917
Integrate Small RNA and Degradome Sequencing to Reveal Drought Memory Response in Wheat (Triticum aestivum L.)
Abstract
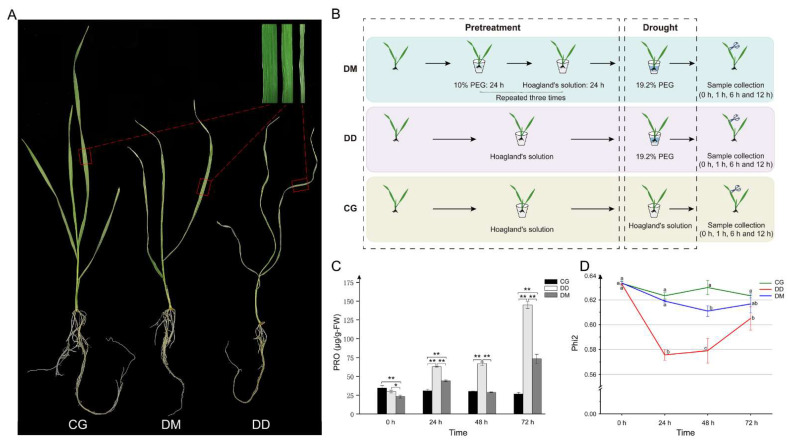
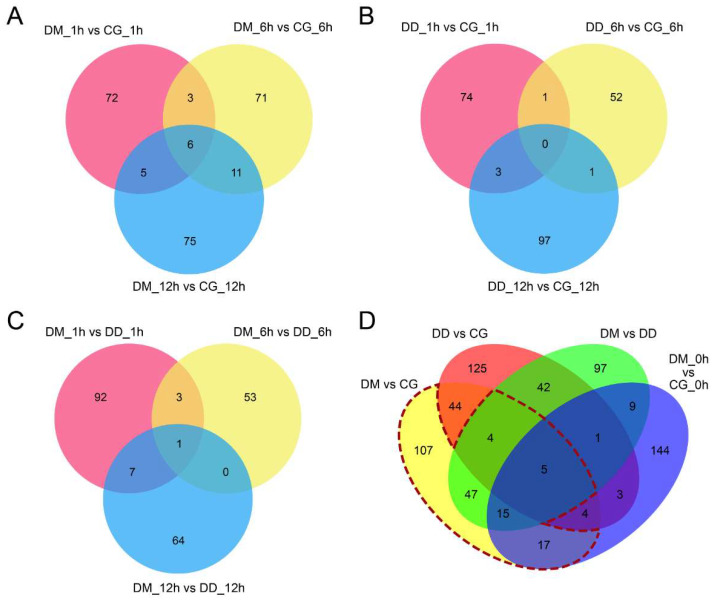
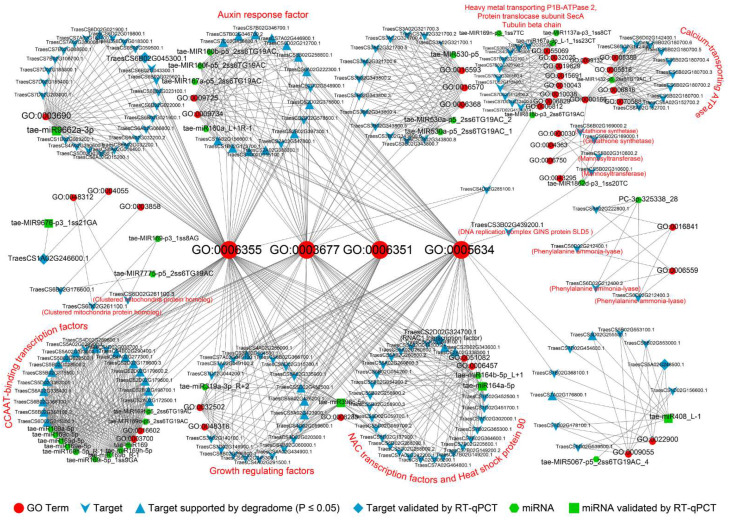
Drought has gradually become one of the most severe abiotic stresses on plants. Plants that experience stress training can exhibit enhanced stress tolerance. According to MicroRNA (miRNA) sequencing data, this study identified 195 candidate drought memory-related miRNAs in wheat, and targets of 64 (32.8%) candidate miRNAs were validated by degradome sequencing. Several drought memory-related miRNAs such as tae-miR9676-5p, tae-MIR9676-p3_1ss21GA, tae-miR171a, tae-miR531_L-2, tae-miR408_L-1, PC-3p-5049_3565, tae-miR396c-5p, tae-miR9778, tae-miR164a-5p, and tae-miR9662a-3p were validated as having a strong response to drought memory by regulating the expression of their target genes. In addition, overexpression of drought memory-related miRNA, tae-miR531_L-2, can remarkably improve the drought tolerance of transgenic Arabidopsisthaliana. Drought memory can regulate plant cellular signal transduction, plant biosynthetic processes, and other biological processes to cope with drought via transcriptional memory. In addition, drought memory-related miRNAs can promote starch and sucrose catabolism and soluble sugar accumulation and regulate proline homeostasis to improve plant drought resistance. Our results could contribute to an understanding of drought memory in wheat seedlings and may provide a new strategy for drought-resistant breeding.
Keywords: drought training; high-throughput sequencing; miR531; proline; transcriptional memory.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



References
-
- Fitton N., Alexander P., Arnell N., Bajzelj B., Calvin K., Doelman J., Gerber J.S., Havlik P., Hasegawa T., Herrero M., et al. The vulnerabilities of agricultural land and food production to future water scarcity. Glob. Environ. Chang.-Hum. Policy Dimens. 2019;58:101944. doi: 10.1016/j.gloenvcha.2019.101944. - DOI
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
