

The antioxidant enzyme Peroxiredoxin-1 controls stroke-associated microglia against acute ischemic stroke
- PMID: 35688114
- PMCID: PMC9184746
- DOI: 10.1016/j.redox.2022.102347
The antioxidant enzyme Peroxiredoxin-1 controls stroke-associated microglia against acute ischemic stroke
Abstract
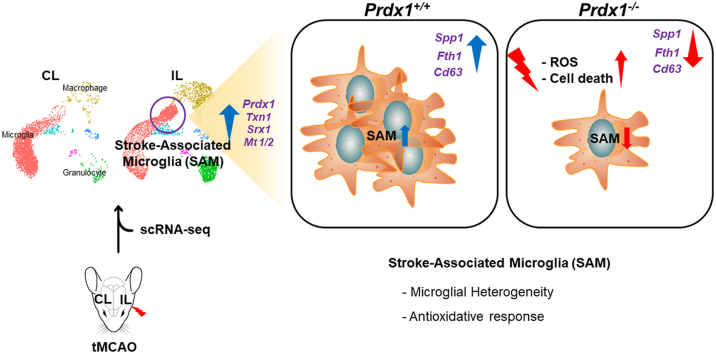
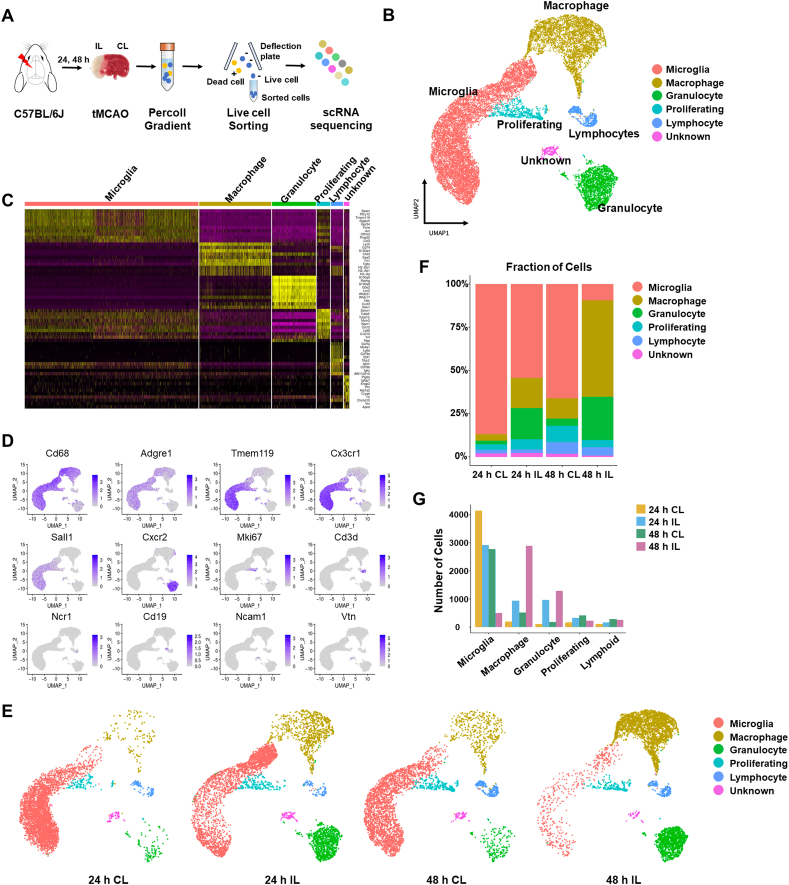
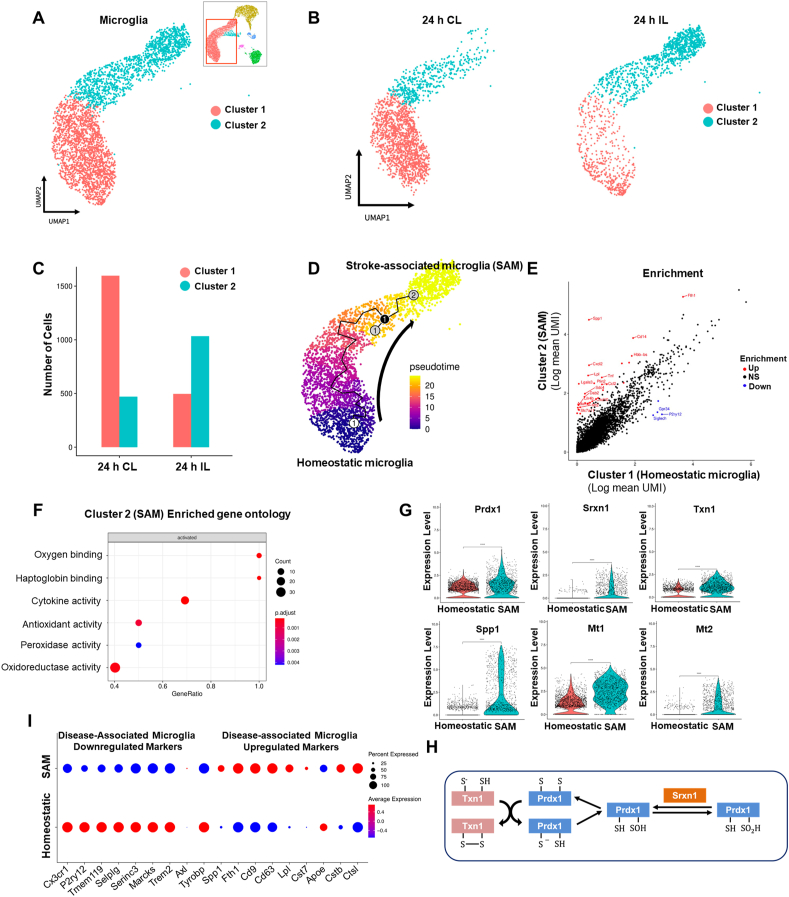
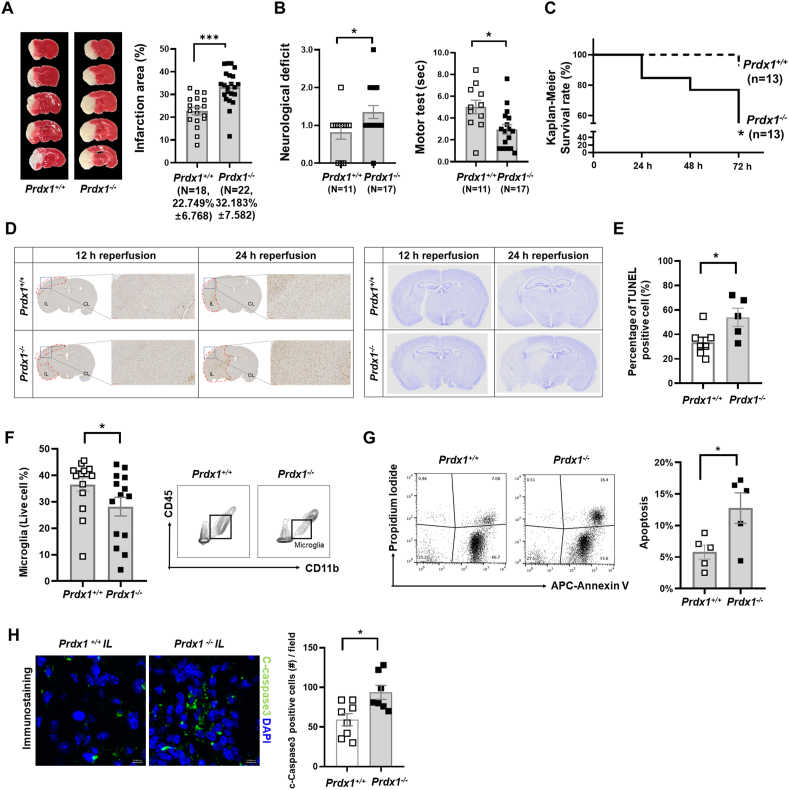
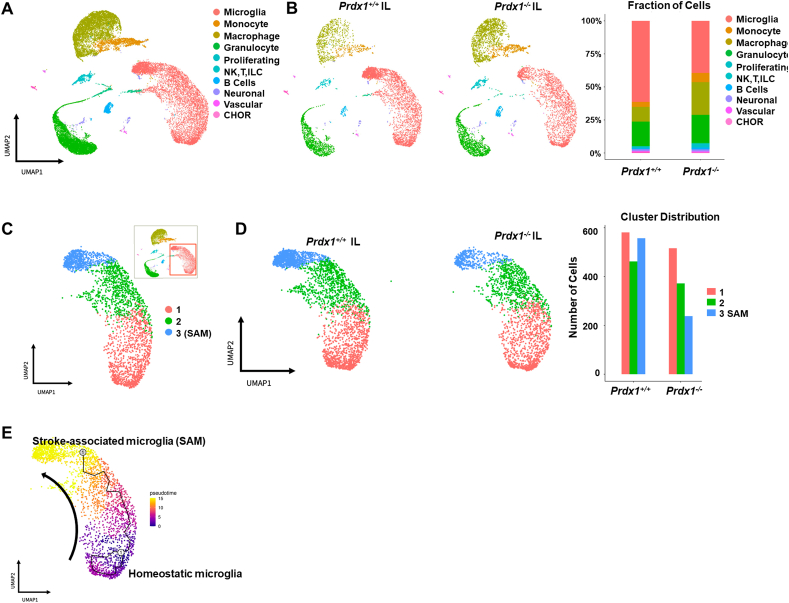
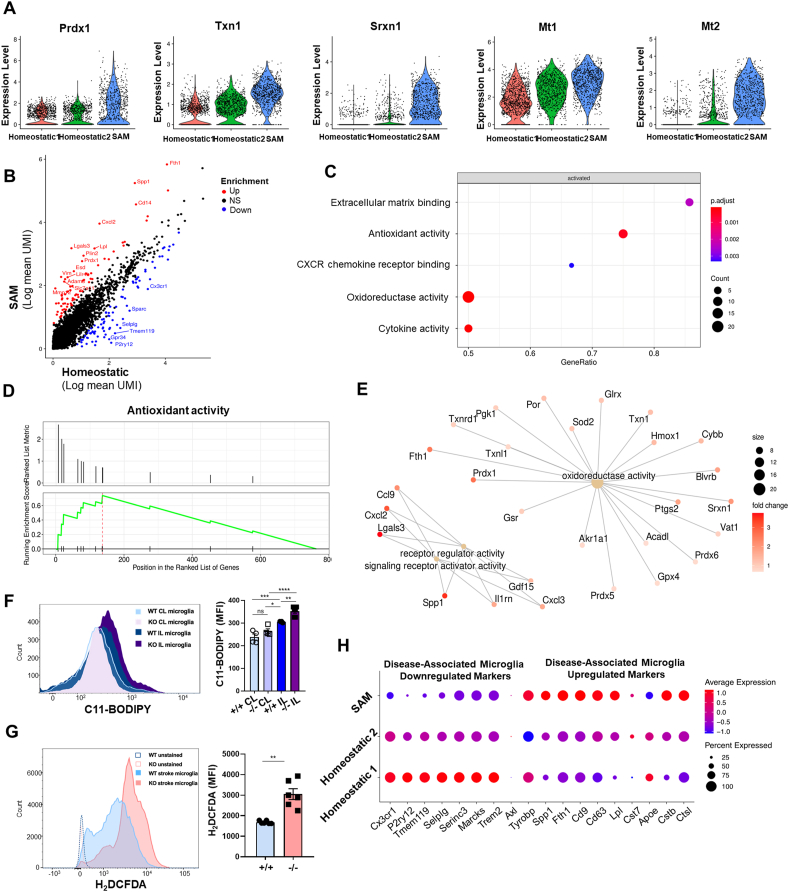
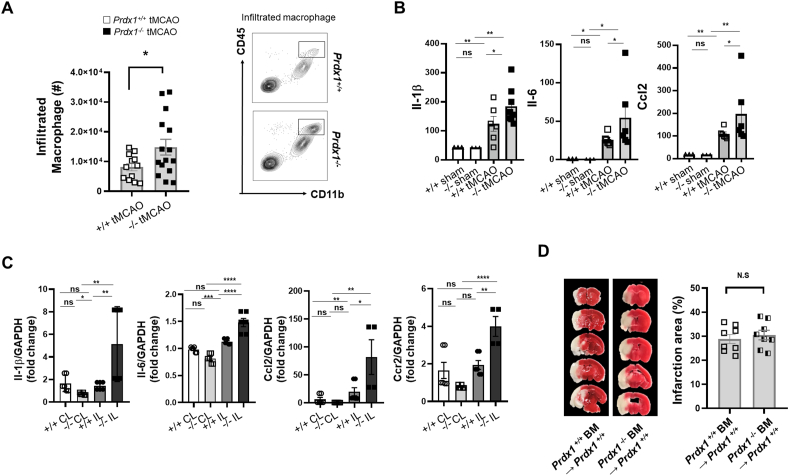
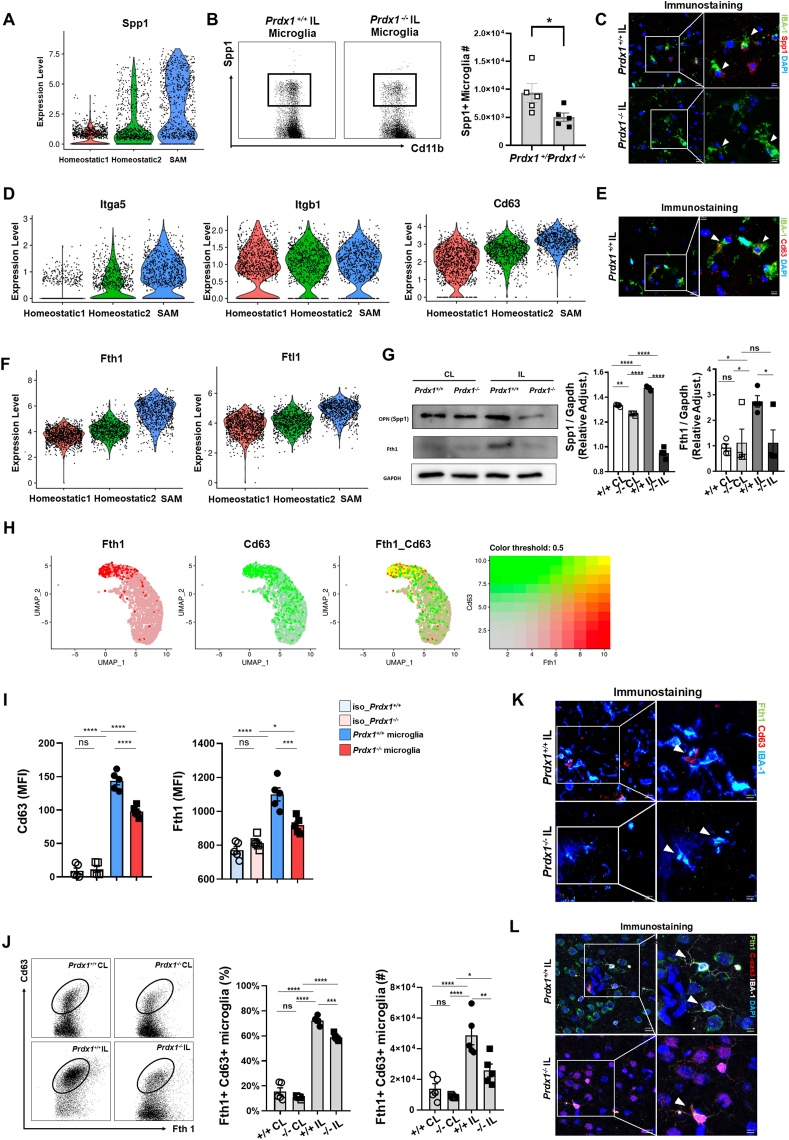
Ischemic stroke is the leading cause of immortal disability and death worldwide. For treatment in the acute phase, it is necessary to control excessive reactive oxygen species (ROS) damage during ischemia/reperfusion (I/R). Microglia are well known to be closely associated with excessive ROS response in the early stage of I/R. However, the precise roles of microglia associated with mitigating ROS damage, and molecular markers of heterogenetic microglia in the I/R damaged brain has not been clarified. Here, we identified a new type of microglia associated with stroke in the I/R injured brain. Single-cell RNA sequencing (scRNA-seq) was used to assess transcriptional changes of microglia and immune cells in the contralateral (CL) and ipsilateral (IL) hemispheres after transient middle cerebral artery occlusion (tMCAO) surgery to mimic ischemic stroke. We classified a unique type of microglia with enhanced antioxidant function and markers similar to those of disease-associated microglia (DAM), designated them as stroke-associated microglia (SAM). The representative antioxidant enzyme, Peroxiredoxin-1 (Prdx1), was predominantly expressed in SAM and mediated ROS defense genes, including Txn1, Srx1, Mt1, and Mt2. In the Prdx1-/- I/R damaged brain, we observed significantly increased infarction, as assessed by TTC staining, and FACS analysis detected severe microglial cell death. Importantly, scRNA transcriptomics data showed that the SAM population was specifically decreased in Prdx1-/- mice and that these mice exhibited decreased ROS damage resistance. Inflammatory responses which were detected by ELISA and qPCR, were also increased in Prdx1-/- IL hemispheres. Finally, Prdx1-dependent antioxidative SAM were found to be essential for increasing the transcription levels of stroke-protective molecules, such as osteopontin and ferritin. A novel microglia type (SAM) is specifically activated in response to stroke I/R injury, and that Prdx1 expression is required for the activation and enhanced antioxidant function of SAM.
Keywords: Ischemic stroke; Peroxiredoxin-1 (Prdx1); Reactive oxygen species (ROS); Single cell RNA sequencing; Stroke-associated microglia (SAM).
Copyright © 2022 The Authors. Published by Elsevier B.V. All rights reserved.
Figures
References
-
- Campbell B.C.V., et al. Ischaemic stroke. Nat. Rev. Dis. Prim. 2019;5(1):70. - PubMed
-
- Allen C.L., Bayraktutan U. Oxidative stress and its role in the pathogenesis of ischaemic stroke. Int. J. Stroke. 2009;4(6):461–470. - PubMed
-
- Jordao M.J.C., et al. Single-cell profiling identifies myeloid cell subsets with distinct fates during neuroinflammation. Science. 2019;363(6425) - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
