

Structure and regulation of the nuclear exosome targeting complex guides RNA substrates to the exosome
- PMID: 35688157
- PMCID: PMC9278407
- DOI: 10.1016/j.molcel.2022.04.011
Structure and regulation of the nuclear exosome targeting complex guides RNA substrates to the exosome
Abstract
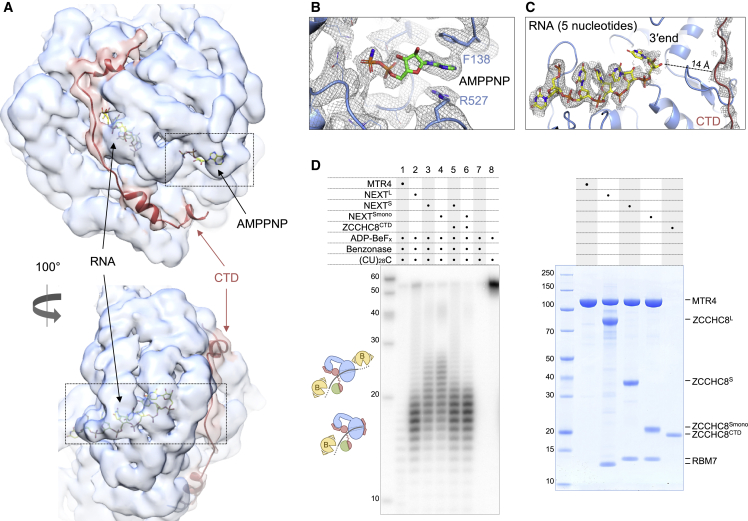
In mammalian cells, spurious transcription results in a vast repertoire of unproductive non-coding RNAs, whose deleterious accumulation is prevented by rapid decay. The nuclear exosome targeting (NEXT) complex plays a central role in directing non-functional transcripts to exosome-mediated degradation, but the structural and molecular mechanisms remain enigmatic. Here, we elucidated the architecture of the human NEXT complex, showing that it exists as a dimer of MTR4-ZCCHC8-RBM7 heterotrimers. Dimerization preconfigures the major MTR4-binding region of ZCCHC8 and arranges the two MTR4 helicases opposite to each other, with each protomer able to function on many types of RNAs. In the inactive state of the complex, the 3' end of an RNA substrate is enclosed in the MTR4 helicase channel by a ZCCHC8 C-terminal gatekeeping domain. The architecture of a NEXT-exosome assembly points to the molecular and regulatory mechanisms with which the NEXT complex guides RNA substrates to the exosome.
Keywords: NEXT; RNA degradation; RNA exosome; RNA processing; conformational regulation; cryo-EM; domain swapping; helicase; non-coding RNAs; pervasive transcription.
Copyright © 2022 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests E.C. is a member of the Molecular Cell advisory board.
Figures






Comment in
-
What's NEXT for the exosome?Mol Cell. 2022 Jul 7;82(13):2355-2357. doi: 10.1016/j.molcel.2022.06.017. Mol Cell. 2022. PMID: 35803215
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
