

Excess ribosomal protein production unbalances translation in a model of Fragile X Syndrome
- PMID: 35688821
- PMCID: PMC9187743
- DOI: 10.1038/s41467-022-30979-0
Excess ribosomal protein production unbalances translation in a model of Fragile X Syndrome
Abstract
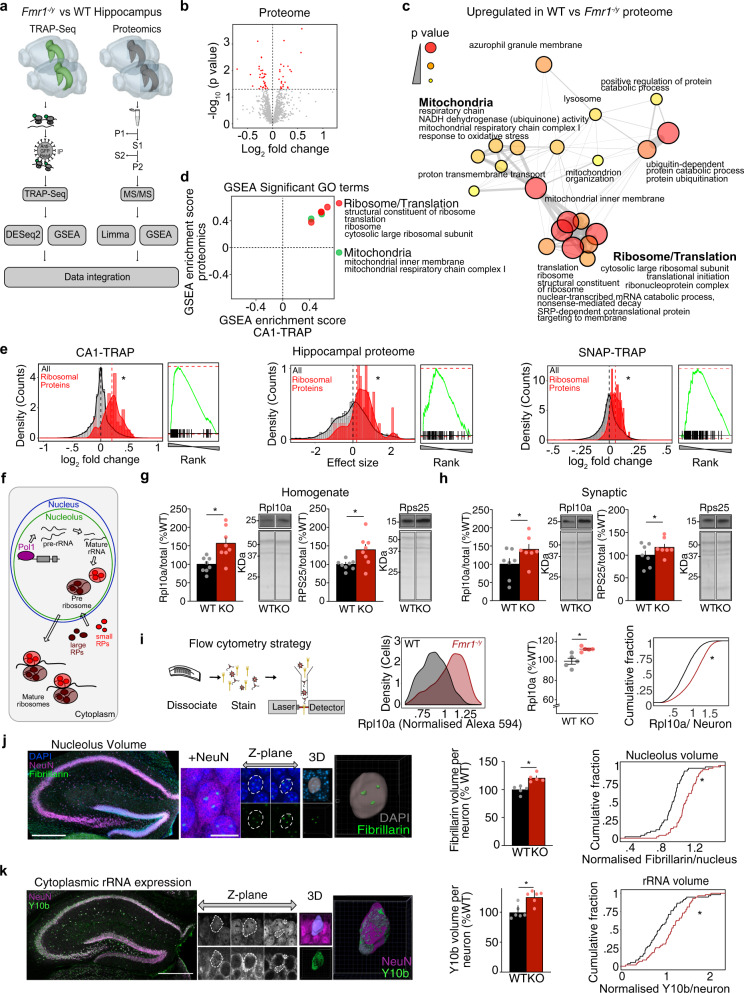
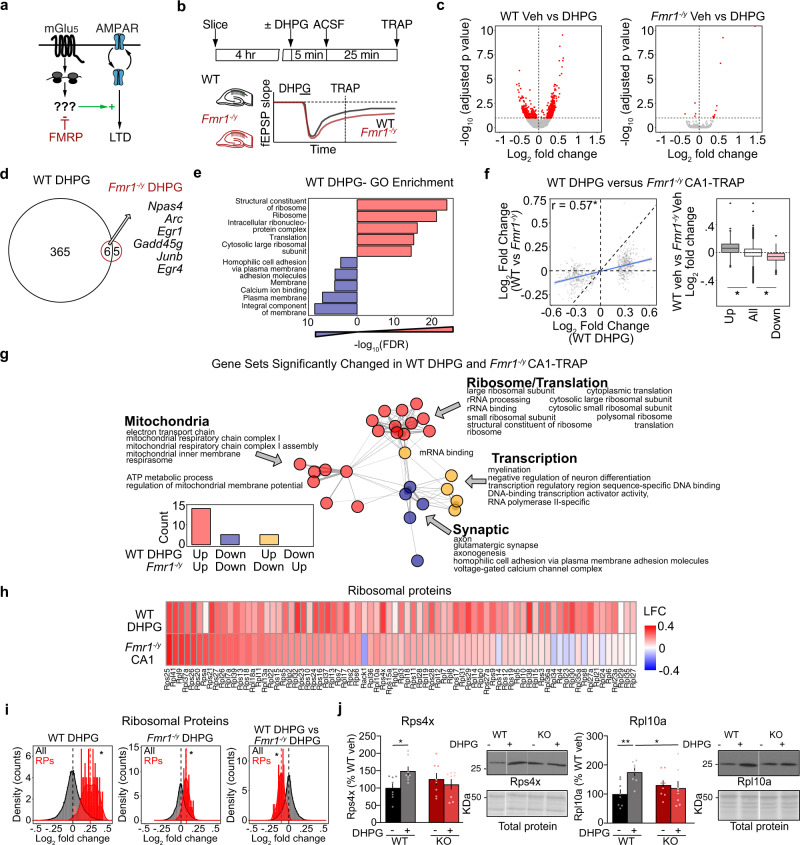
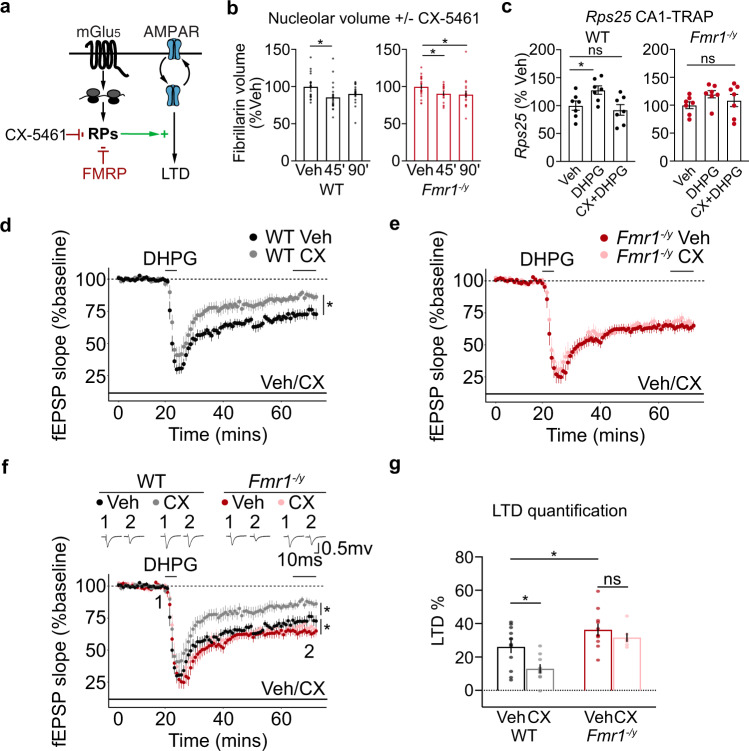
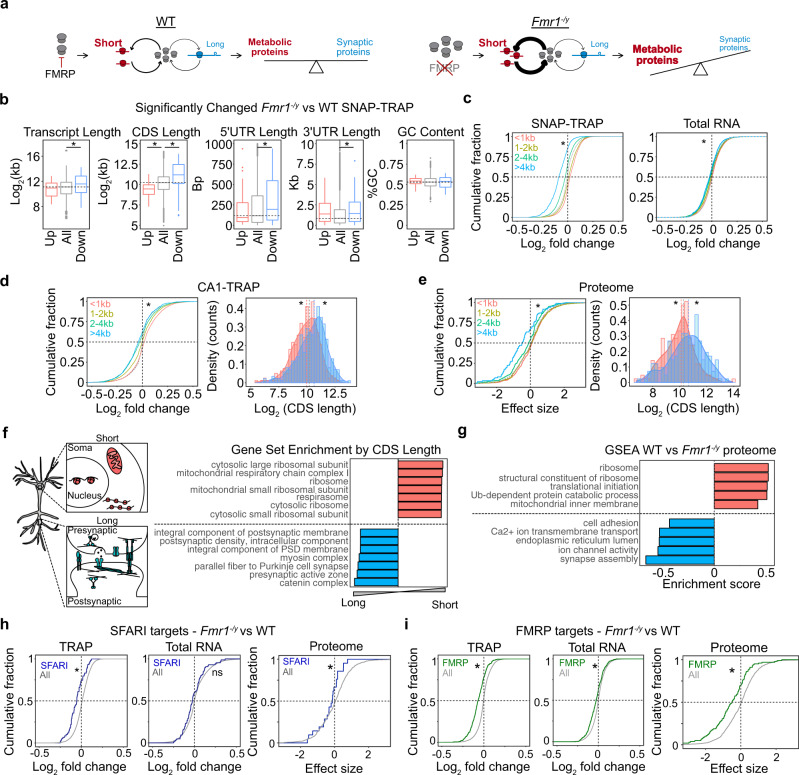
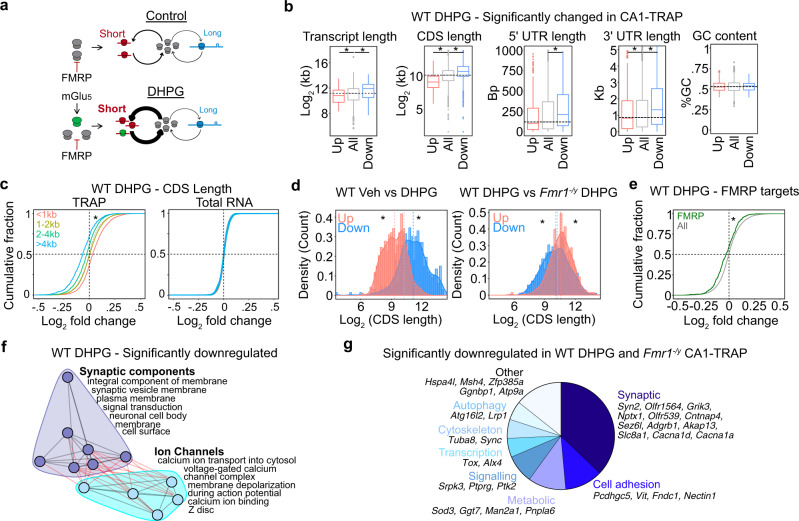
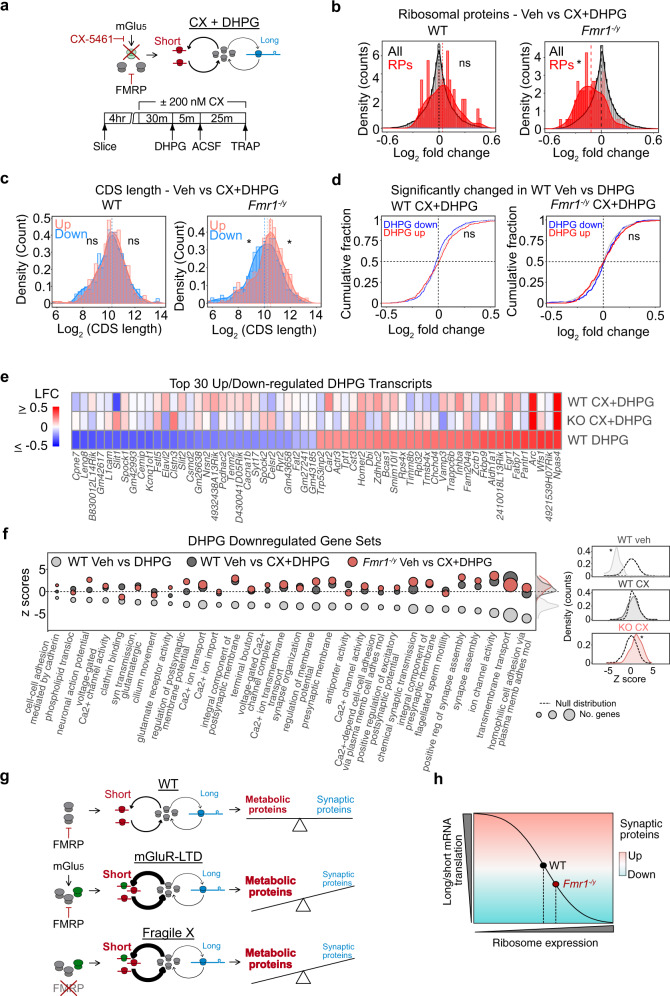
Dysregulated protein synthesis is a core pathogenic mechanism in Fragile X Syndrome (FX). The mGluR Theory of FX predicts that pathological synaptic changes arise from the excessive translation of mRNAs downstream of mGlu1/5 activation. Here, we use a combination of CA1 pyramidal neuron-specific TRAP-seq and proteomics to identify the overtranslating mRNAs supporting exaggerated mGlu1/5 -induced long-term synaptic depression (mGluR-LTD) in the FX mouse model (Fmr1-/y). Our results identify a significant increase in the translation of ribosomal proteins (RPs) upon mGlu1/5 stimulation that coincides with a reduced translation of long mRNAs encoding synaptic proteins. These changes are mimicked and occluded in Fmr1-/y neurons. Inhibiting RP translation significantly impairs mGluR-LTD and prevents the length-dependent shift in the translating population. Together, these results suggest that pathological changes in FX result from a length-dependent alteration in the translating population that is supported by excessive RP translation.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
