

A juvenile locomotor program promotes vocal learning in zebra finches
- PMID: 35689094
- PMCID: PMC9187677
- DOI: 10.1038/s42003-022-03533-3
A juvenile locomotor program promotes vocal learning in zebra finches
Abstract
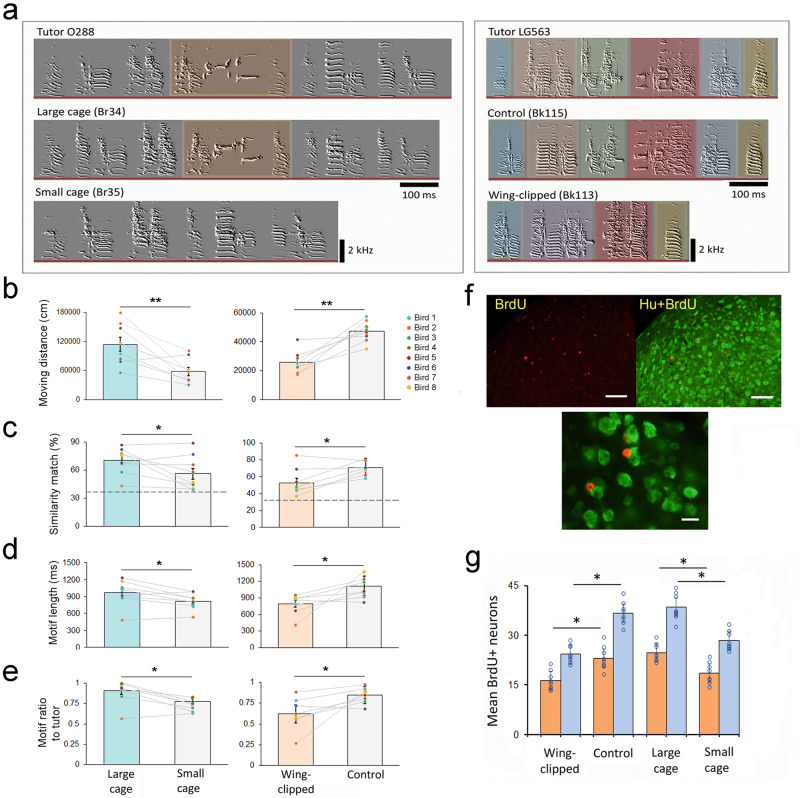
The evolution and development of complex, learned motor skills are thought to be closely associated with other locomotor movement and cognitive functions. However, it remains largely unknown how different neuromuscular programs may interconnect during the protracted developmental process. Here we use a songbird to examine the behavioral and neural substrates between the development of locomotor movement and vocal-motor learning. Juvenile songbirds escalate their locomotor activity during the sensitive period for vocal learning, followed by a surge of vocal practice. Individual variability of locomotor production is positively correlated with precision of tutor imitation and duration of multi-syllable sequences. Manipulation of juvenile locomotion significantly impacts the precision of vocal imitation and neural plasticity. The locomotor program developed during the sensitive period of vocal learning may enrich the neural substrates that promote the subsequent development of vocal learning.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures



References
-
- Allott, R. The motor theory of language: origin and function. 105–119 (1992).
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
