

Processes and mechanisms of coastal woody-plant mortality
- PMID: 35689431
- PMCID: PMC9544010
- DOI: 10.1111/gcb.16297
Processes and mechanisms of coastal woody-plant mortality
Abstract
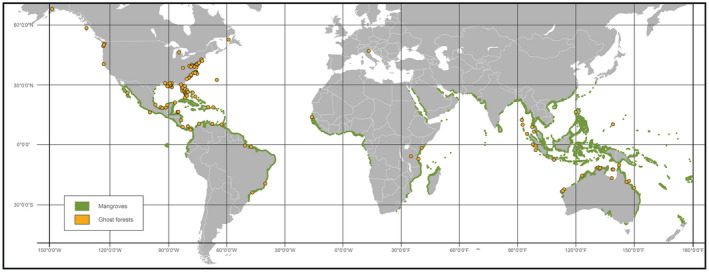
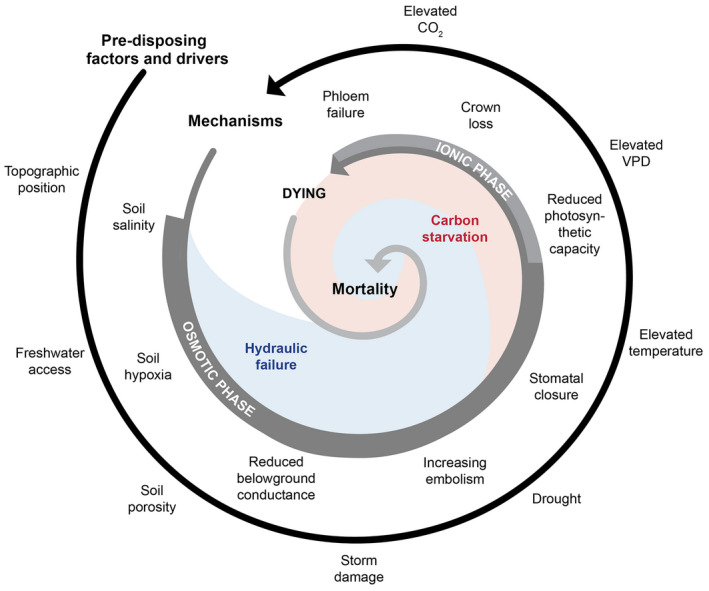
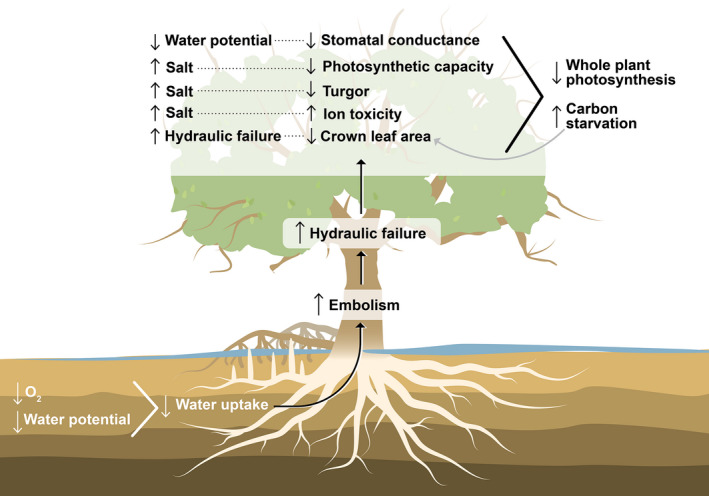
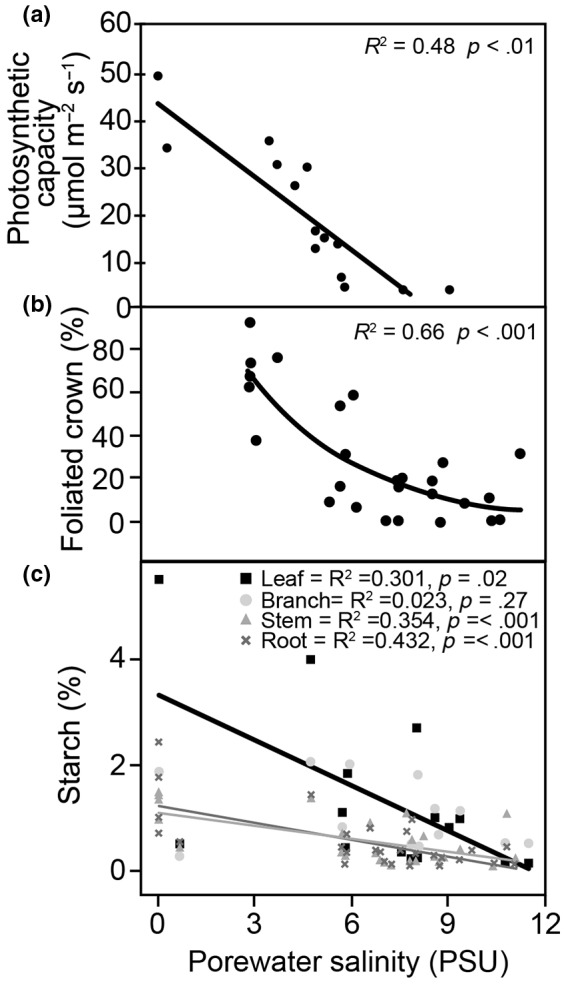
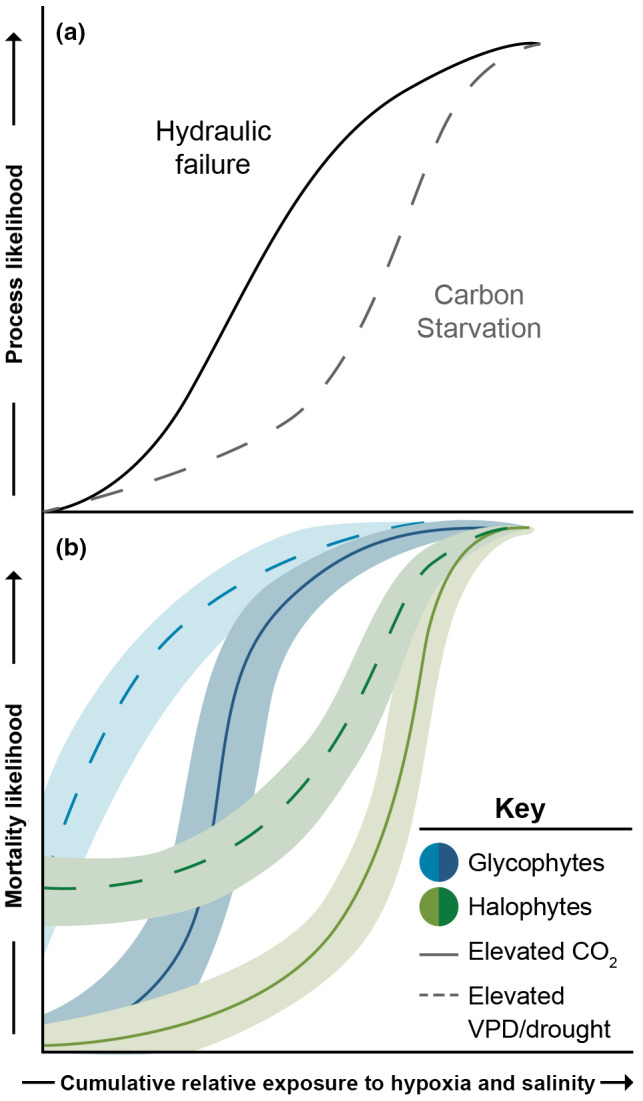
Observations of woody plant mortality in coastal ecosystems are globally widespread, but the overarching processes and underlying mechanisms are poorly understood. This knowledge deficiency, combined with rapidly changing water levels, storm surges, atmospheric CO2 , and vapor pressure deficit, creates large predictive uncertainty regarding how coastal ecosystems will respond to global change. Here, we synthesize the literature on the mechanisms that underlie coastal woody-plant mortality, with the goal of producing a testable hypothesis framework. The key emergent mechanisms underlying mortality include hypoxic, osmotic, and ionic-driven reductions in whole-plant hydraulic conductance and photosynthesis that ultimately drive the coupled processes of hydraulic failure and carbon starvation. The relative importance of these processes in driving mortality, their order of progression, and their degree of coupling depends on the characteristics of the anomalous water exposure, on topographic effects, and on taxa-specific variation in traits and trait acclimation. Greater inundation exposure could accelerate mortality globally; however, the interaction of changing inundation exposure with elevated CO2 , drought, and rising vapor pressure deficit could influence mortality likelihood. Models of coastal forests that incorporate the frequency and duration of inundation, the role of climatic drivers, and the processes of hydraulic failure and carbon starvation can yield improved estimates of inundation-induced woody-plant mortality.
Keywords: carbon starvation; climate change; coastal; hydraulic failure; hypoxia; mortality; salinity; sea level rise.
© 2022 Battelle Memorial Institute and The Authors. Global Change Biology published by John Wiley & Sons Ltd. This article has been contributed to by U.S. Government employees and their work is in the public domain in the USA.
Figures

References
-
- Acosta‐Motos, J. R. , Ortuño, M. F. , Bernal‐Vicente, A. , Diaz‐Vivancos, P. , Sanchez‐Blanco, M. J. , & Hernandez, J. A. (2017). Plant responses to salt stress: Adaptive mechanisms. Agronomy, 7(1), 18.
-
- Adams, H. D. , Zeppel, M. J. , Anderegg, W. R. , Hartmann, H. , Landhäusser, S. M. , Tissue, D. T. , Huxman, T. E. , Hudson, P. J. , Franz, T. E. , Allen, C. D. , & Anderegg, L. D. (2017). A multi‐species synthesis of physiological mechanisms in drought‐induced tree mortality. Nature Ecology & Evolution, 1(9), 1285–1291. - PubMed
-
- Ainsworth, E. A. , & Rogers, A. (2007). The response of photosynthesis and stomatal conductance to rising [CO2]: Mechanisms and environmental interactions. Plant, Cell & Environment, 30(3), 258–270. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
