

Neural consequences of chronic sleep disruption
- PMID: 35691776
- PMCID: PMC9388586
- DOI: 10.1016/j.tins.2022.05.007
Neural consequences of chronic sleep disruption
Abstract
Recent studies in both humans and animal models call into question the completeness of recovery after chronic sleep disruption. Studies in humans have identified cognitive domains particularly vulnerable to delayed or incomplete recovery after chronic sleep disruption, including sustained vigilance and episodic memory. These findings, in turn, provide a focus for animal model studies to critically test the lasting impact of sleep loss on the brain. Here, we summarize the human response to sleep disruption and then discuss recent findings in animal models examining recovery responses in circuits pertinent to vigilance and memory. We then propose pathways of injury common to various forms of sleep disruption and consider the implications of this injury in aging and in neurodegenerative disorders.
Keywords: amyloid; locus coeruleus; neurodegeneration; sleep deprivation; sleep fragmentation; sleep restriction.
Copyright © 2022 The Author(s). Published by Elsevier Ltd.. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
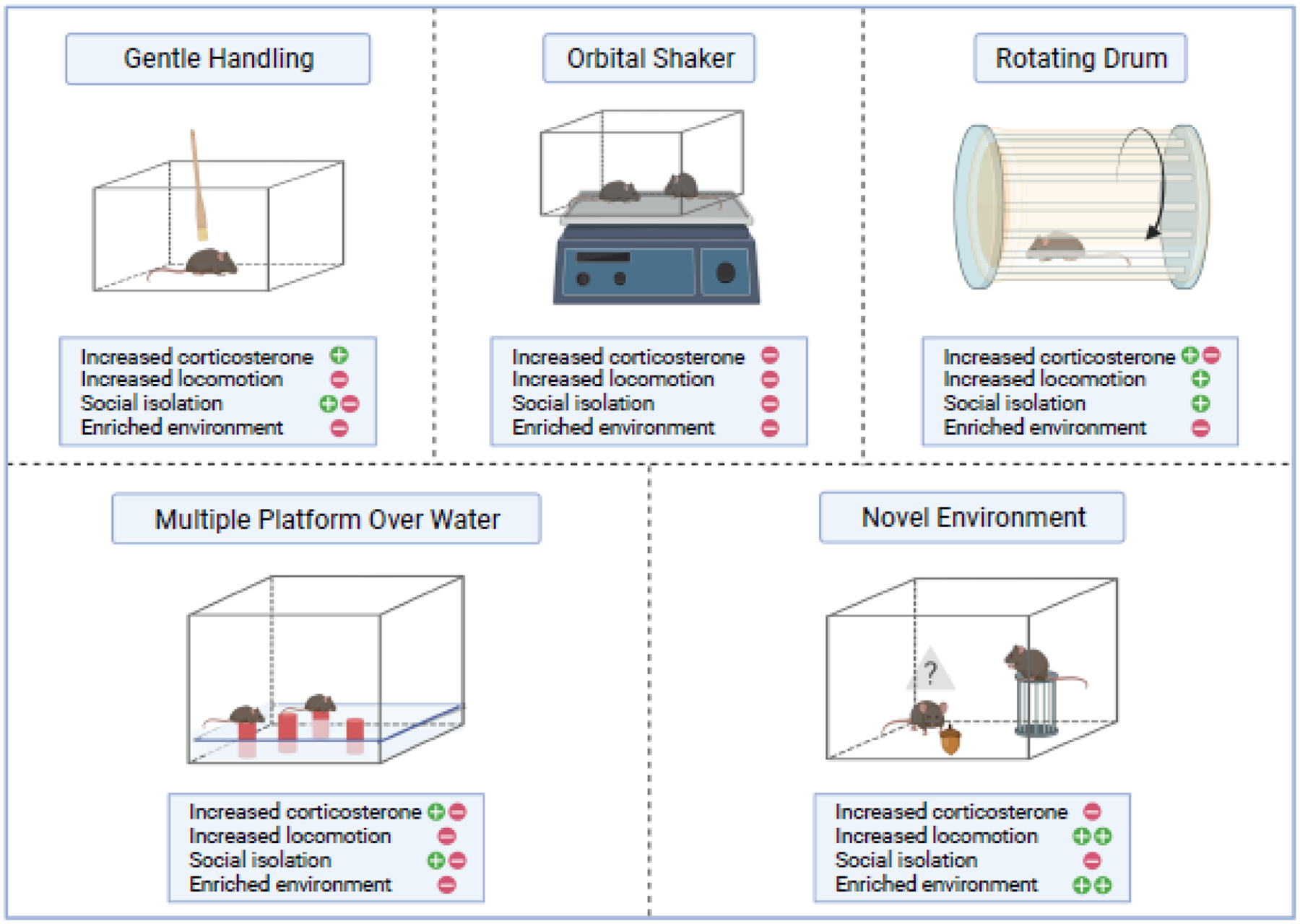

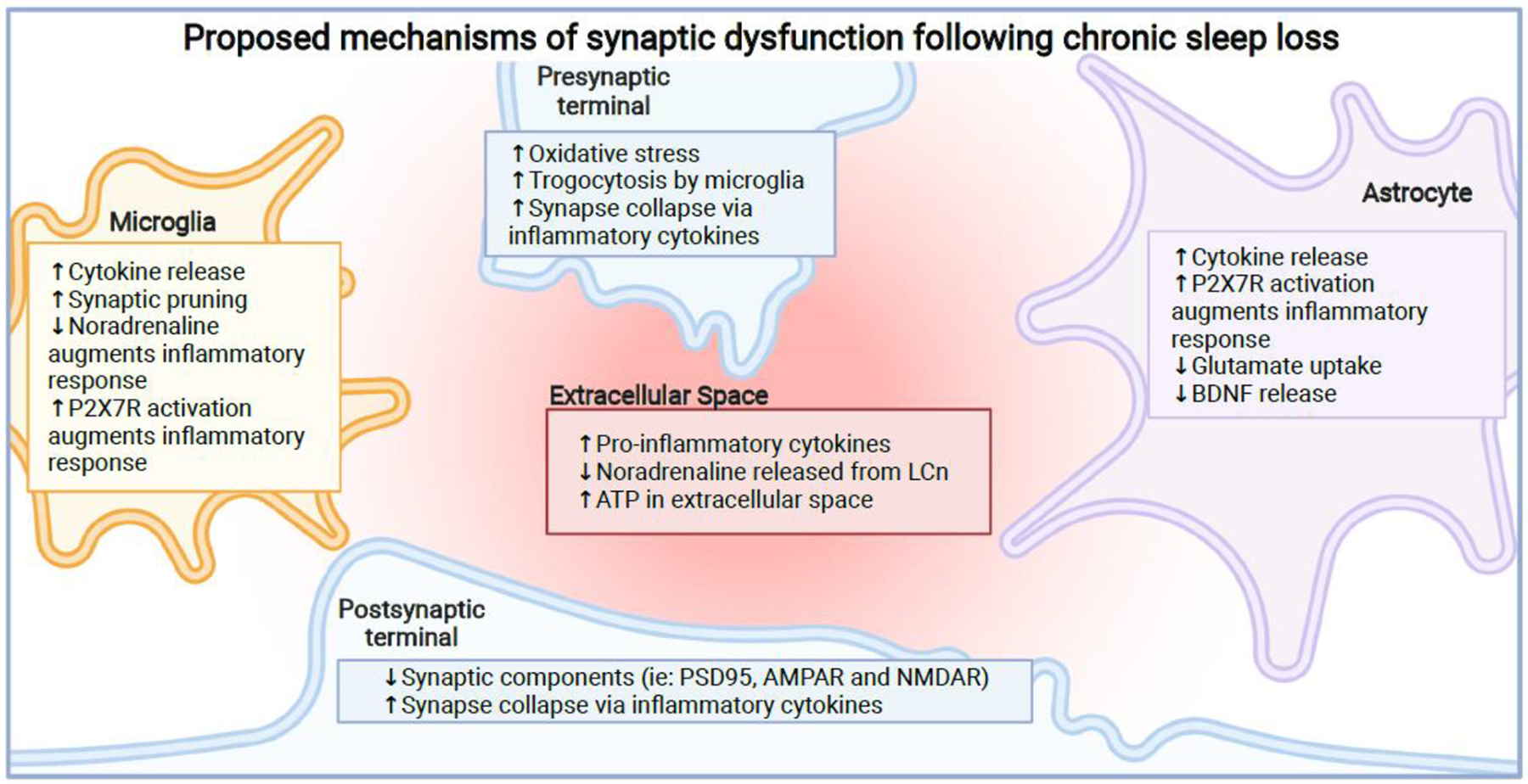
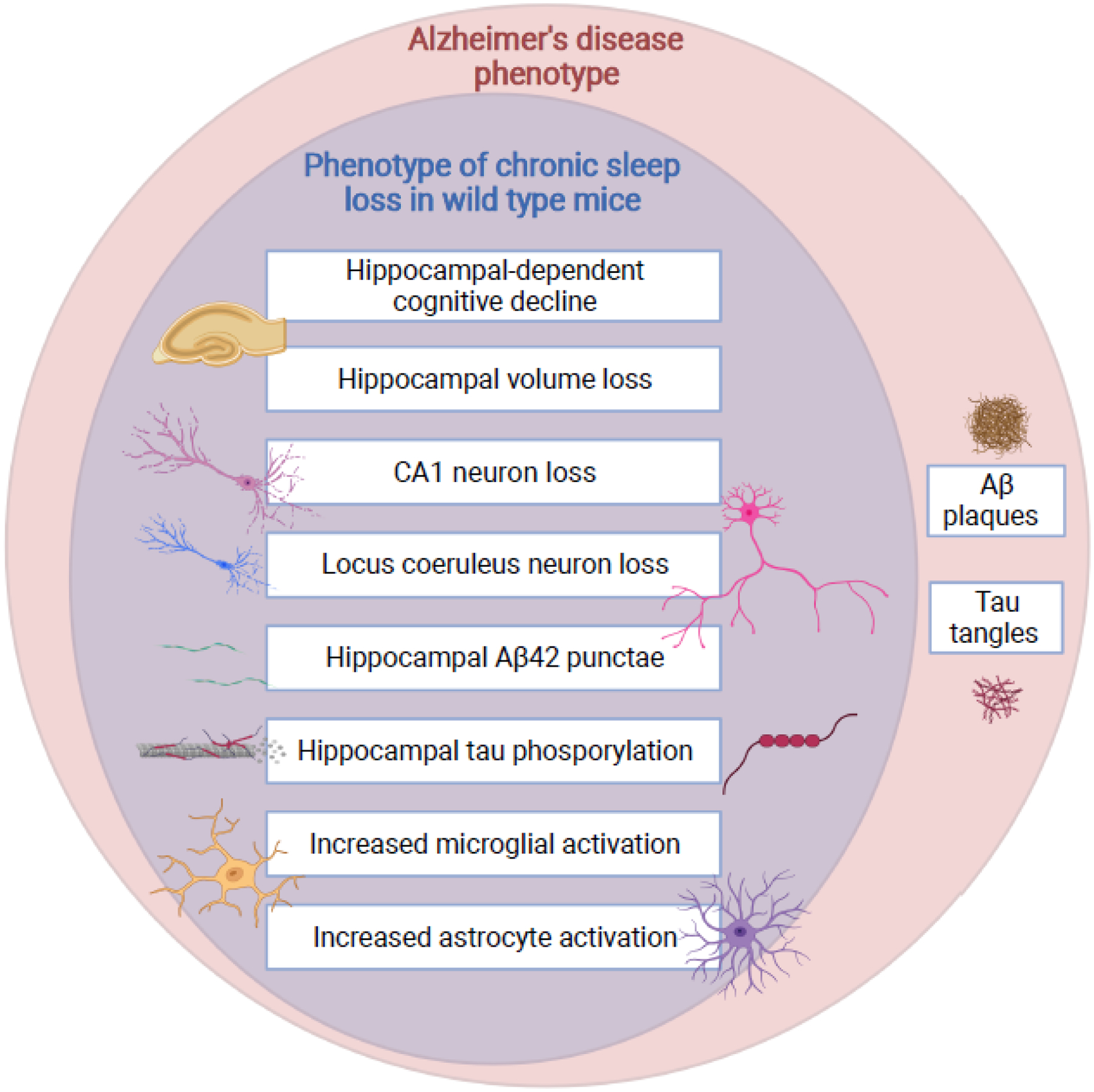
References
-
- Dinges DF et al. (1997) Cumulative sleepiness, mood disturbance, and psychomotor vigilance performance decrements during a week of sleep restricted to 4–5 hours per night. Sleep 20, 267–277 - PubMed
-
- Van Dongen HP et al. (2003) The cumulative cost of additional wakefulness: dose-response effects on neurobehavioral functions and sleep physiology from chronic sleep restriction and total sleep deprivation. Sleep 26, 117–126 - PubMed
-
- Belenky G et al. (2003) Patterns of performance degradation and restoration during sleep restriction and subsequent recovery: a sleep dose-response study. J Sleep Res 12, 1–12. 337 [pii] - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
