

Novel Small Molecule Tyrosine Kinase 2 Pseudokinase Ligands Block Cytokine-Induced TYK2-Mediated Signaling Pathways
- PMID: 35693820
- PMCID: PMC9186491
- DOI: 10.3389/fimmu.2022.884399
Novel Small Molecule Tyrosine Kinase 2 Pseudokinase Ligands Block Cytokine-Induced TYK2-Mediated Signaling Pathways
Abstract
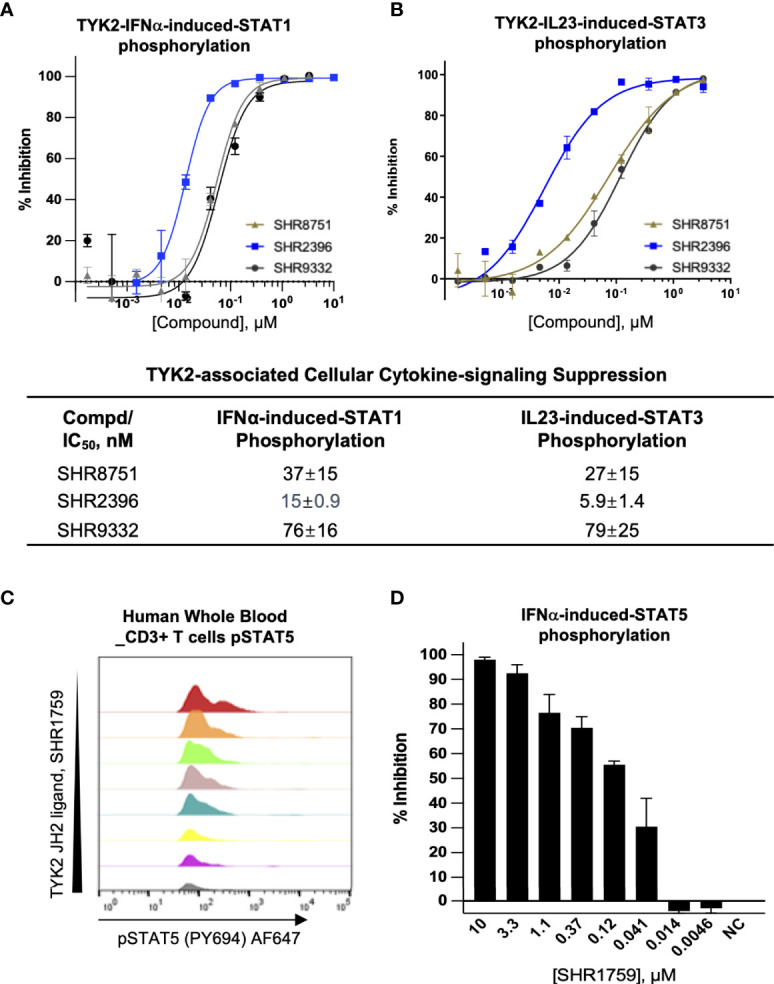
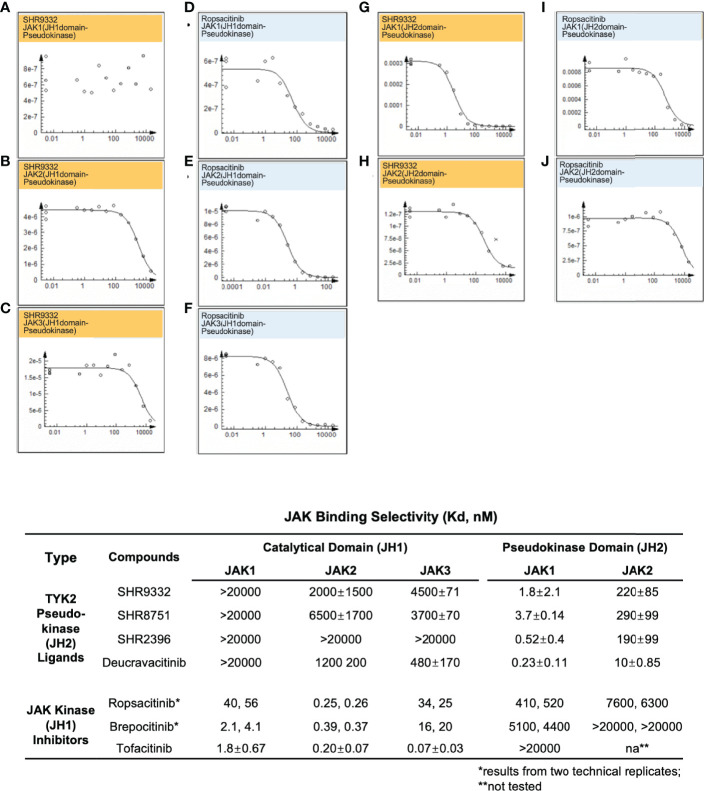
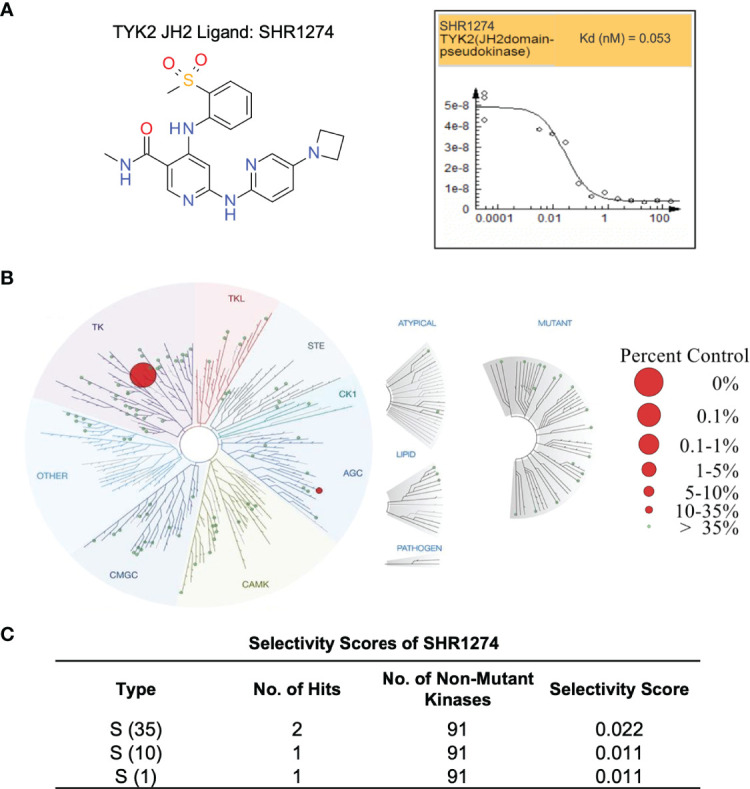
A member of the Janus kinase (JAK) family, Tyrosine Kinase 2 (TYK2), is crucial in mediating various cytokine-signaling pathways such as interleukin-23 (IL23), interleukin-12 (IL12) and type I Interferons (IFN) which contribute to autoimmune disorders (e.g., psoriasis, lupus, and inflammatory bowel disease). Thus, TYK2 represents an attractive target to develop small-molecule therapeutics for the treatment of cytokine-driven inflammatory diseases. Selective inhibition of TYK2 over other JAK isoforms is critical to achieve a favorable therapeutic index in the development of TYK2 inhibitors. However, designing small molecule inhibitors to target the adenosine triphosphate (ATP) binding site of TYK2 kinase has been challenging due to the substantial structural homology of the JAK family catalytic domains. Here, we employed an approach to target the JAK homology 2 (JH2) pseudokinase regulatory domain of the TYK2 protein. We developed a series of small-molecule TYK2 pseudokinase ligands, which suppress the TYK2 catalytic activity through allosteric regulation. The TYK2 pseudokinase-binding small molecules in this study simultaneously achieve high affinity-binding for the TYK2 JH2 domain while also affording significantly reduced affinity for the TYK2 JAK homology 1 (JH1) kinase domain. These TYK2 JH2 selective molecules, although possessing little effect on suppressing the catalytic activity of the isolated TYK2 JH1 catalytic domain in the kinase assays, can still significantly block the TYK2-mediated receptor-stimulated pathways by binding to the TYK2 JH2 domain and allosterically regulating the TYK2 JH1 kinase. These compounds are potent towards human T-cell lines and primary immune cells as well as in human whole-blood specimens. Moreover, TYK2 JH2-binding ligands exhibit remarkable selectivity of TYK2 over JAK isoforms not only biochemically but also in a panel of receptor-stimulated JAK1/JAK2/JAK3-driven cellular functional assays. In addition, the TYK2 JH2-targeting ligands also demonstrate high selectivity in a multi-kinase screening panel. The data in the current study underscores that the TYK2 JH2 pseudokinase is a promising therapeutic target for achieving a high degree of biological selectivity. Meanwhile, targeting the JH2 domain represents an appealing strategy for the development of clinically well-tolerated TYK2 inhibitors that would have superior efficacy and a favorable safety profile compared to the existing Janus kinase inhibitors against autoimmune diseases.
Keywords: JAK homology 2 (JH2) pseudokinase regulatory domain; Janus Kinases (JAK); Tyrosine Kinase 2 (TYK2); cytokine pathways; selectivity; small molecule inhibitors.
Copyright © 2022 Zhou, Li, Shen, Wang, Zhang, Liu, Li, Liu, Li, Yan, Dong, Zhang, Wu, Zhuang, Chowdhury, Miller, Issa, Mao, Chen, Feng, Li, Bai, He and Tao.
Conflict of interest statement
YZ, RS, XW, FZ, DL, JLiu, SL, YY, HW, JLi, PL, LZ, RC, MM, and MI were employees of Eternity Bioscience Inc. XL, PD, ZZ, YM, JF, HC, CB, FH, and WT are employees and shareholders of Hengrui Pharmaceutical Co. Ltd.
Figures




Similar articles
-
A novel highly selective allosteric inhibitor of tyrosine kinase 2 (TYK2) can block inflammation- and autoimmune-related pathways.Cell Commun Signal. 2023 Oct 16;21(1):287. doi: 10.1186/s12964-023-01299-7. Cell Commun Signal. 2023. PMID: 37845748 Free PMC article.
-
Structural and Functional Characterization of the JH2 Pseudokinase Domain of JAK Family Tyrosine Kinase 2 (TYK2).J Biol Chem. 2015 Nov 6;290(45):27261-27270. doi: 10.1074/jbc.M115.672048. Epub 2015 Sep 10. J Biol Chem. 2015. PMID: 26359499 Free PMC article.
-
Deucravacitinib is an allosteric TYK2 protein kinase inhibitor FDA-approved for the treatment of psoriasis.Pharmacol Res. 2023 Mar;189:106642. doi: 10.1016/j.phrs.2022.106642. Epub 2023 Feb 6. Pharmacol Res. 2023. PMID: 36754102 Review.
-
The pseudokinase domain is required for suppression of basal activity of Jak2 and Jak3 tyrosine kinases and for cytokine-inducible activation of signal transduction.J Biol Chem. 2002 Dec 6;277(49):47954-63. doi: 10.1074/jbc.M205156200. Epub 2002 Sep 25. J Biol Chem. 2002. PMID: 12351625
-
Tyk2 Targeting in Immune-Mediated Inflammatory Diseases.Int J Mol Sci. 2023 Feb 8;24(4):3391. doi: 10.3390/ijms24043391. Int J Mol Sci. 2023. PMID: 36834806 Free PMC article. Review.
Cited by
-
High-Resolution Structure of RNA G-Quadruplex Containing Unique Structural Motifs Originating from the 5'-UTR of Human Tyrosine Kinase 2 (TYK2).ACS Omega. 2024 Feb 2;9(6):7215-7229. doi: 10.1021/acsomega.3c09592. eCollection 2024 Feb 13. ACS Omega. 2024. PMID: 38371751 Free PMC article.
-
Update on the pathophysiology and treatment of primary Sjögren syndrome.Nat Rev Rheumatol. 2024 Aug;20(8):473-491. doi: 10.1038/s41584-024-01135-3. Epub 2024 Jul 9. Nat Rev Rheumatol. 2024. PMID: 38982205 Review.
-
Biomarkers Associated with Drugs for the Treatment of Lupus Nephritis.Biomolecules. 2023 Oct 31;13(11):1601. doi: 10.3390/biom13111601. Biomolecules. 2023. PMID: 38002282 Free PMC article. Review.
-
Mechanisms and therapeutic prospect of the JAK-STAT signaling pathway in liver cancer.Mol Cell Biochem. 2025 Jan;480(1):1-17. doi: 10.1007/s11010-024-04983-5. Epub 2024 Mar 22. Mol Cell Biochem. 2025. PMID: 38519710 Review.
-
Deucravacitinib, a tyrosine kinase 2 pseudokinase inhibitor, protects human EndoC-βH1 β-cells against proinflammatory insults.Front Immunol. 2023 Oct 3;14:1263926. doi: 10.3389/fimmu.2023.1263926. eCollection 2023. Front Immunol. 2023. PMID: 37854597 Free PMC article.
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous