

Unbalanced selection: the challenge of maintaining a social polymorphism when a supergene is selfish
- PMID: 35694754
- PMCID: PMC9189496
- DOI: 10.1098/rstb.2021.0197
Unbalanced selection: the challenge of maintaining a social polymorphism when a supergene is selfish
Abstract
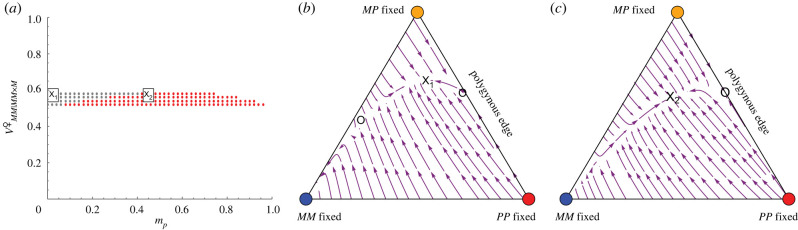
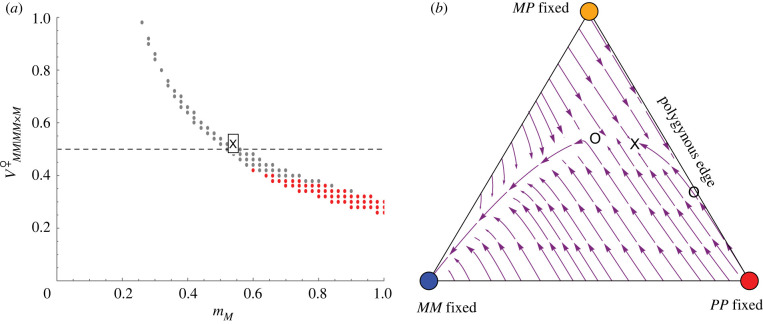
Supergenes often have multiple phenotypic effects, including unexpected detrimental ones, because recombination suppression maintains associations among co-adapted alleles but also allows the accumulation of recessive deleterious mutations and selfish genetic elements. Yet, supergenes often persist over long evolutionary periods. How are such polymorphisms maintained in the face of selection, drive and drift? We present a population genetic model that investigates the conditions necessary for a stable polymorphic equilibrium when one of the supergene haplotypes is a selfish genetic element. The model fits the characteristics of the Alpine silver ant, Formica selysi, in which a large supergene underlies colony social organization, and one haplotype distorts Mendelian transmission by killing progeny that did not inherit it. The model shows that such maternal-effect killing strongly limits the maintenance of social polymorphism. Under random mating, transmission ratio distortion prevents rare single-queen colonies from invading populations of multiple-queen colonies, regardless of the fitness of each genotype. A stable polymorphic equilibrium can, however, be reached when high rates of assortative mating are combined with large fitness differences among supergene genotypes. The model reveals that the persistence of the social polymorphism is non-trivial and expected to occur only under restrictive conditions that deserve further empirical investigation. This article is part of the theme issue 'Genomic architecture of supergenes: causes and evolutionary consequences'.
Keywords: ants; genetic polymorphism; heterozygote advantage; invasion analysis; selfish genetic element; social organization.
Conflict of interest statement
We declare we have no competing interests.
Figures
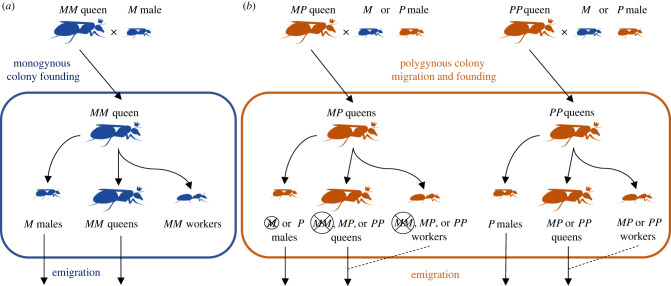
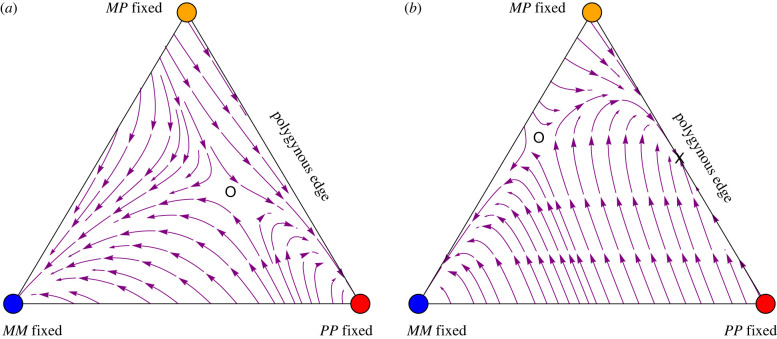
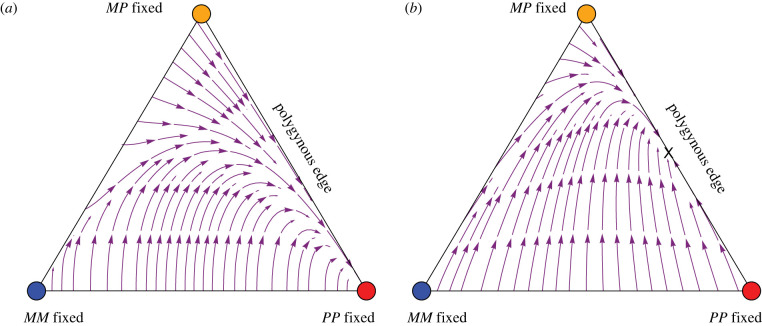
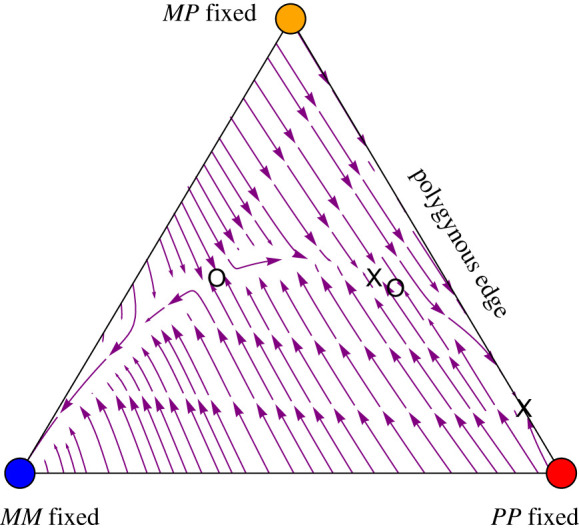
References
-
- Dobzhansky T. 1970. Genetics of the evolutionary process. New York, NY: Columbia University Press.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources