

Evolutionary origin and functional diversification of aminotransferases
- PMID: 35697072
- PMCID: PMC9309667
- DOI: 10.1016/j.jbc.2022.102122
Evolutionary origin and functional diversification of aminotransferases
Abstract
Aminotransferases (ATs) are pyridoxal 5'-phosphate-dependent enzymes that catalyze the transamination reactions between amino acid donor and keto acid acceptor substrates. Modern AT enzymes constitute ∼2% of all classified enzymatic activities, play central roles in nitrogen metabolism, and generate multitude of primary and secondary metabolites. ATs likely diverged into four distinct AT classes before the appearance of the last universal common ancestor and further expanded to a large and diverse enzyme family. Although the AT family underwent an extensive functional specialization, many AT enzymes retained considerable substrate promiscuity and multifunctionality because of their inherent mechanistic, structural, and functional constraints. This review summarizes the evolutionary history, diverse metabolic roles, reaction mechanisms, and structure-function relationships of the AT family enzymes, with a special emphasis on their substrate promiscuity and multifunctionality. Comprehensive characterization of AT substrate specificity is still needed to reveal their true metabolic functions in interconnecting various branches of the nitrogen metabolic network in different organisms.
Keywords: PLP-dependent enzymes; amino acids; aminotransferases; enzyme evolution; nitrogen metabolism; transaminases.
Copyright © 2022 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures





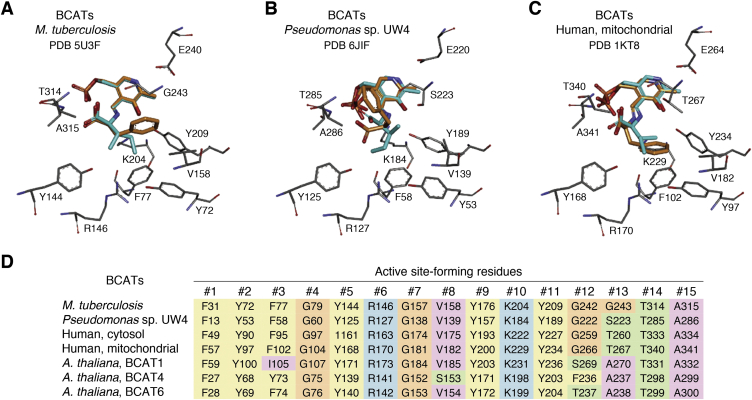
References
-
- Givan C.V. In: Miflin B.J., editor. 329–357. Academic Press; Cambridge, MA: 1980. 8 - aminotransferases in higher plants. (Amino Acids and Derivatives).
-
- Mehta P.K., Hale T.I., Christen P. Aminotransferases: demonstration of homology and division into evolutionary subgroups. Eur. J. Biochem. 1993;214:549–561. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
