

GluN3A excitatory glycine receptors control adult cortical and amygdalar circuits
- PMID: 35700736
- PMCID: PMC9365314
- DOI: 10.1016/j.neuron.2022.05.016
GluN3A excitatory glycine receptors control adult cortical and amygdalar circuits
Abstract
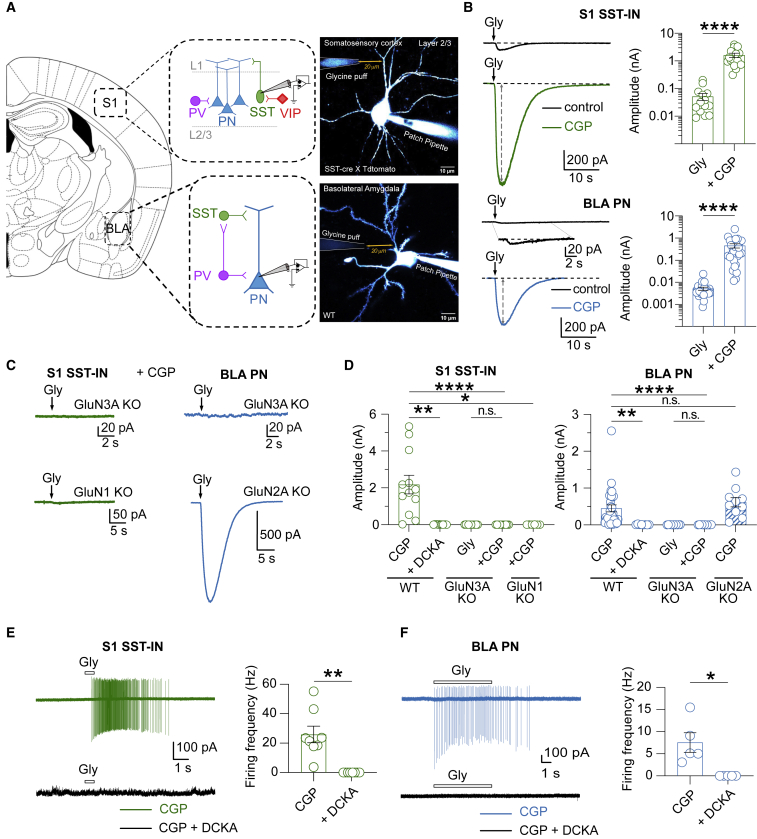
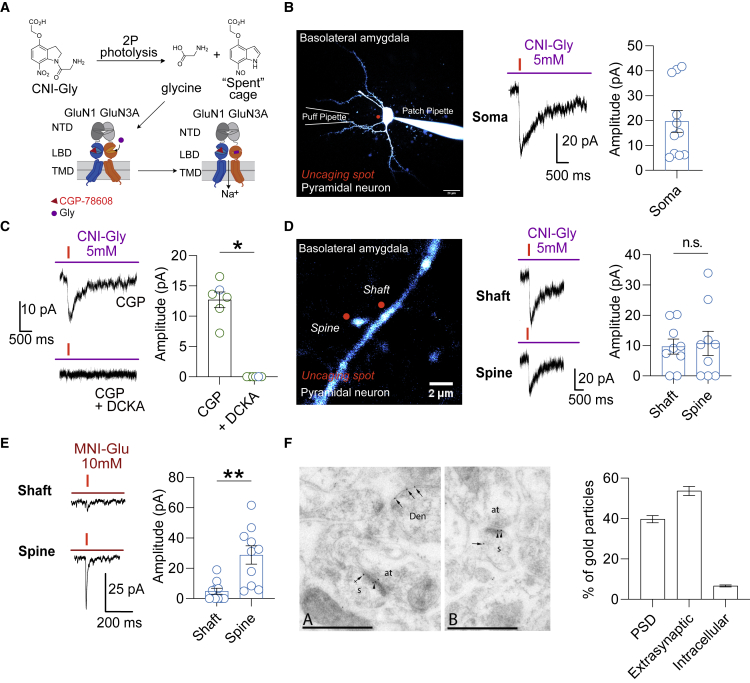
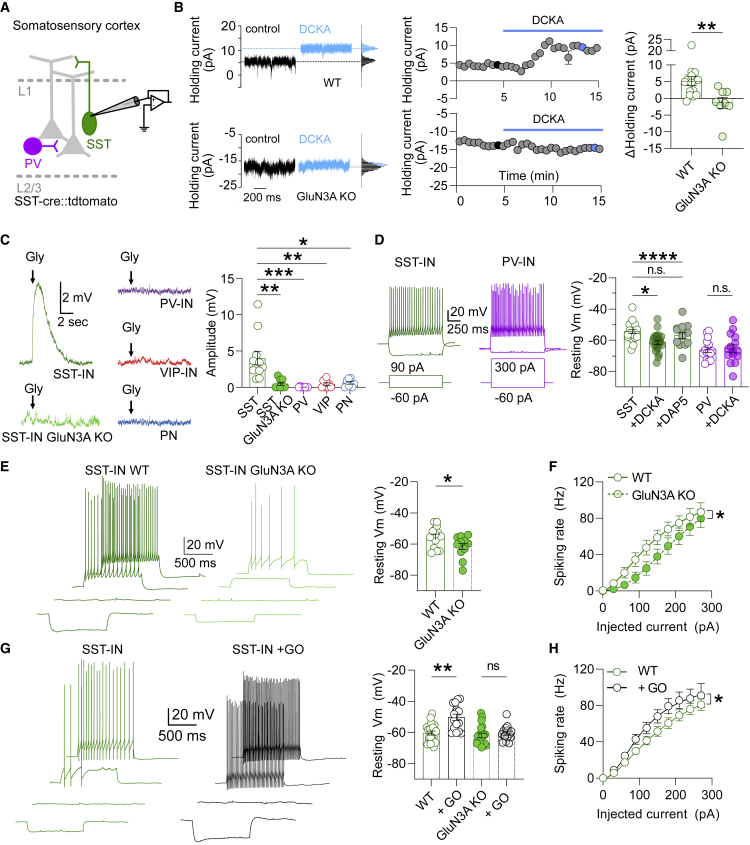
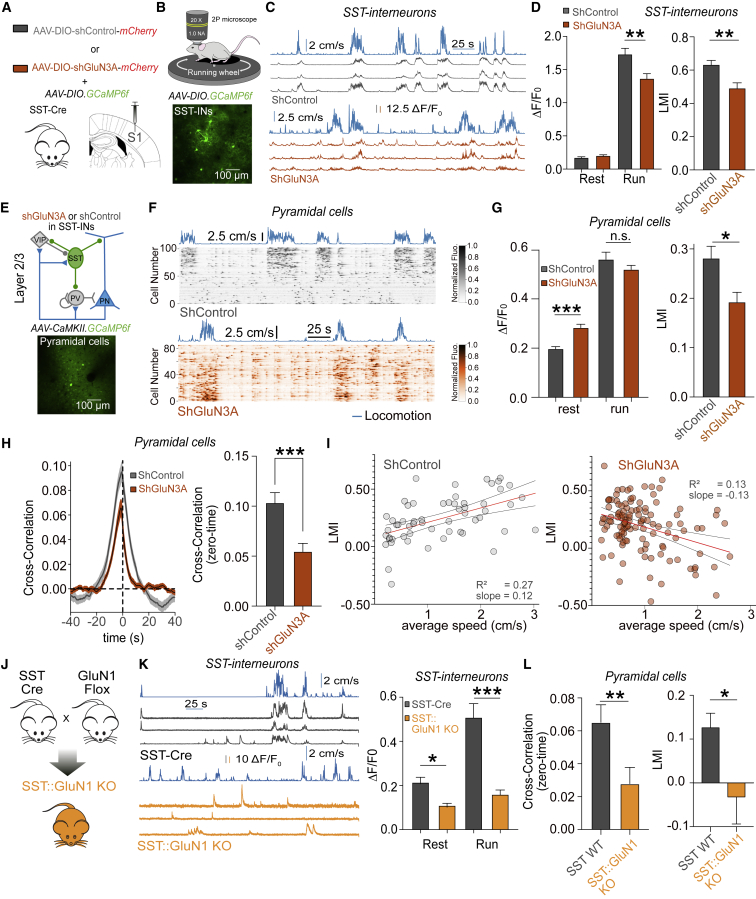
GluN3A is an atypical glycine-binding subunit of NMDA receptors (NMDARs) whose actions in the brain are mostly unknown. Here, we show that the expression of GluN3A subunits controls the excitability of mouse adult cortical and amygdalar circuits via an unusual signaling mechanism involving the formation of excitatory glycine GluN1/GluN3A receptors (eGlyRs) and their tonic activation by extracellular glycine. eGlyRs are mostly extrasynaptic and reside in specific neuronal populations, including the principal cells of the basolateral amygdala (BLA) and SST-positive interneurons (SST-INs) of the neocortex. In the BLA, tonic eGlyR currents are sensitive to fear-conditioning protocols, are subject to neuromodulation by the dopaminergic system, and control the stability of fear memories. In the neocortex, eGlyRs control the in vivo spiking of SST-INs and the behavior-dependent modulation of cortical activity. GluN3A-containing eGlyRs thus represent a novel and widespread signaling modality in the adult brain, with attributes that strikingly depart from those of conventional NMDARs.
Keywords: GluN3A; NMDA; amygdala; cortex; fear; glycine; interneuron; neurotransmission; receptors; somatostatin interneurons; tonic activation.
Copyright © 2022 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures




Comment in
-
The exciting side of unconventional glycine receptors.Neuron. 2022 Aug 3;110(15):2359-2361. doi: 10.1016/j.neuron.2022.07.007. Neuron. 2022. PMID: 35926450
References
-
- Basu A.C., Tsai G.E., Ma C.-L., Ehmsen J.T., Mustafa A.K., Han L., Jiang Z.I., Benneyworth M.A., Froimowitz M.P., Lange N., et al. Targeted disruption of serine racemase affects glutamatergic neurotransmission and behavior. Mol. Psychiatry. 2009;14:719–727. doi: 10.1038/mp.2008.130. - DOI - PMC - PubMed
-
- Ciabarra M., Sevarino A., Gahn G., Pecht G., Heinemann S., Sevarino K.A. Cloning and characterization of chi-1: a developmentally regulated member of a novel class of the ionotropic glutamate receptor family. J Neurosci. 1995;15:6498–6508. doi: 10.1523/JNEUROSCI.15-10-06498.1995. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
