

Morphological pseudotime ordering and fate mapping reveal diversification of cerebellar inhibitory interneurons
- PMID: 35701402
- PMCID: PMC9197879
- DOI: 10.1038/s41467-022-30977-2
Morphological pseudotime ordering and fate mapping reveal diversification of cerebellar inhibitory interneurons
Abstract
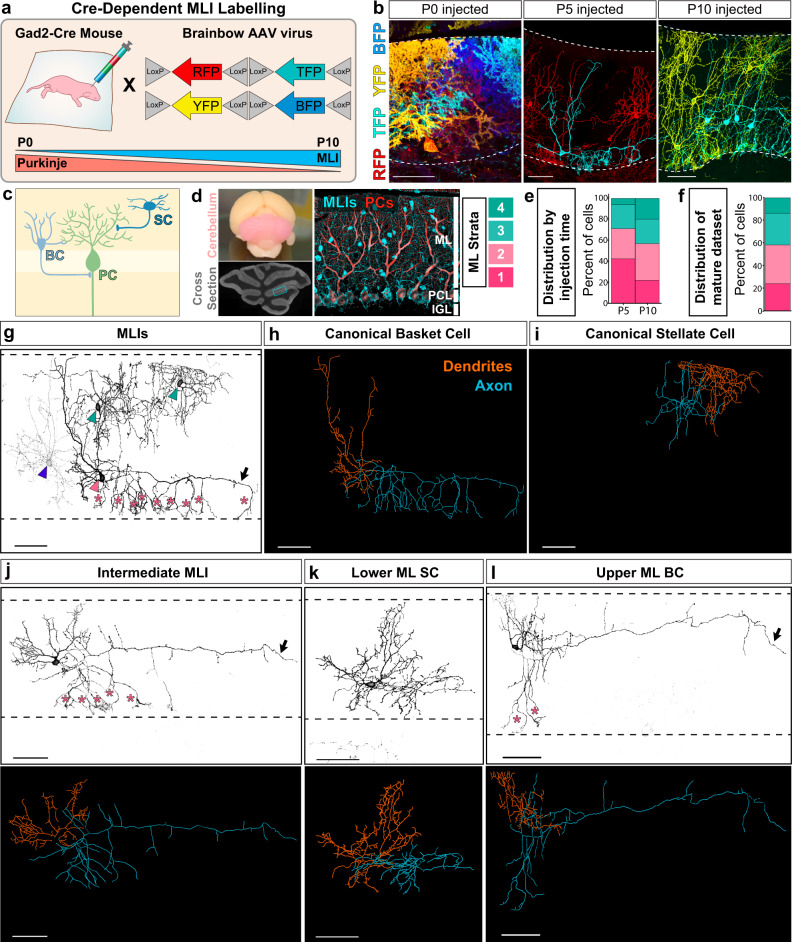
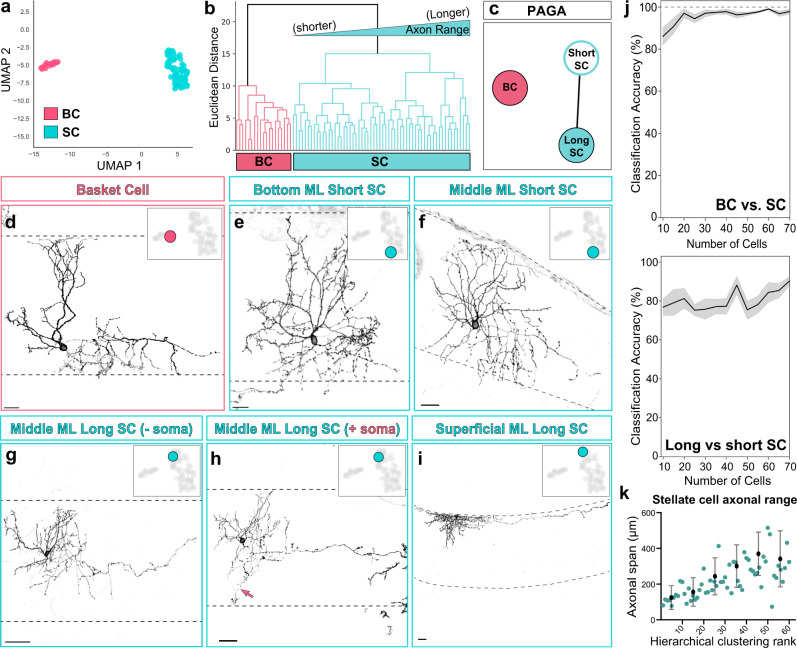
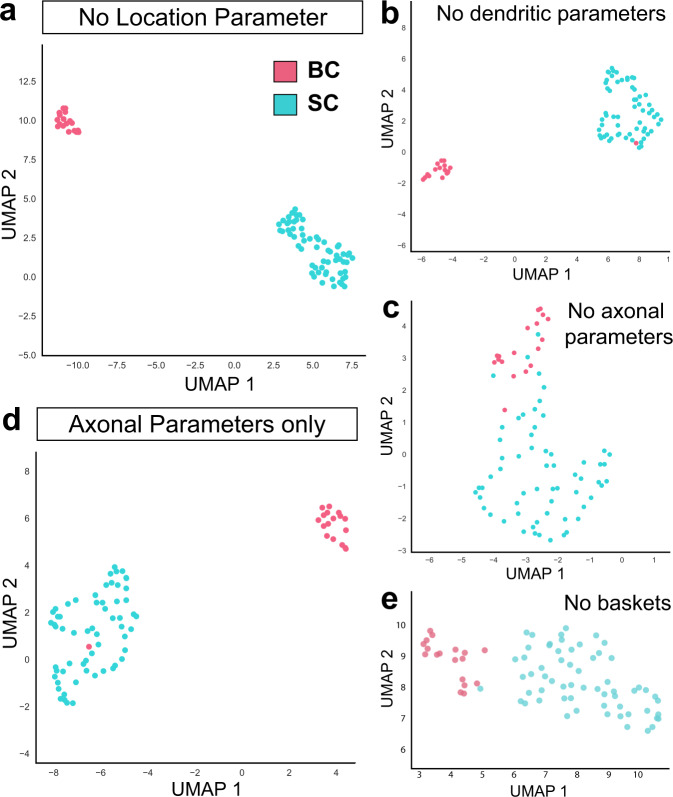
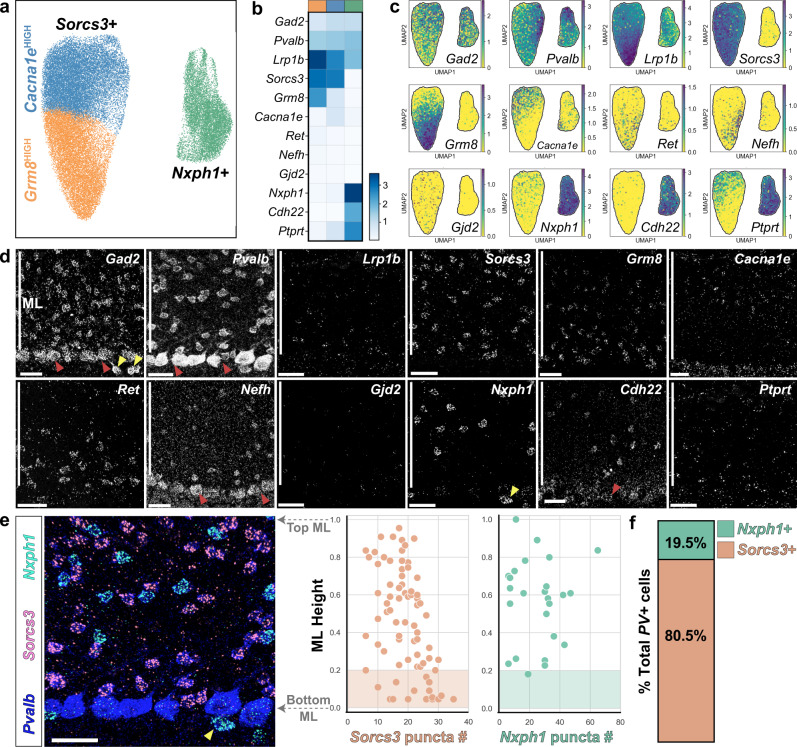
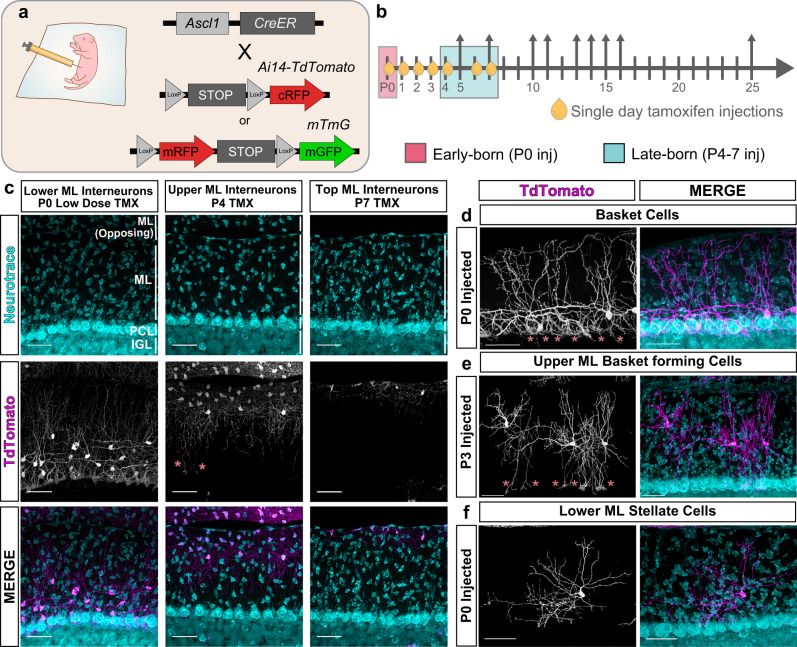
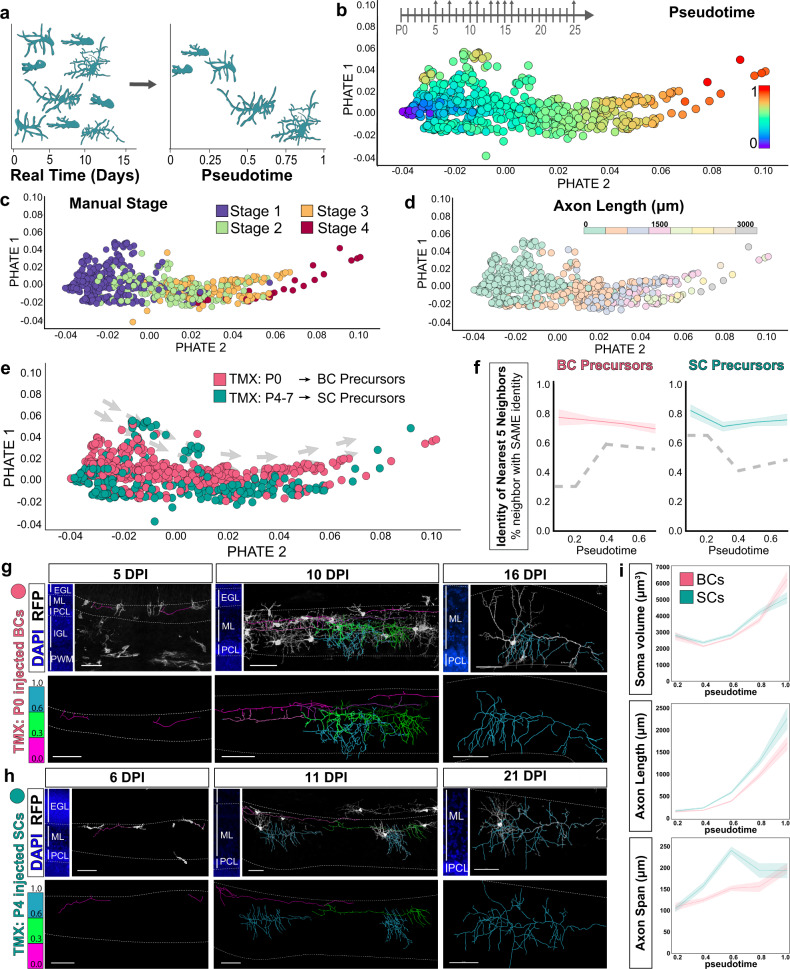
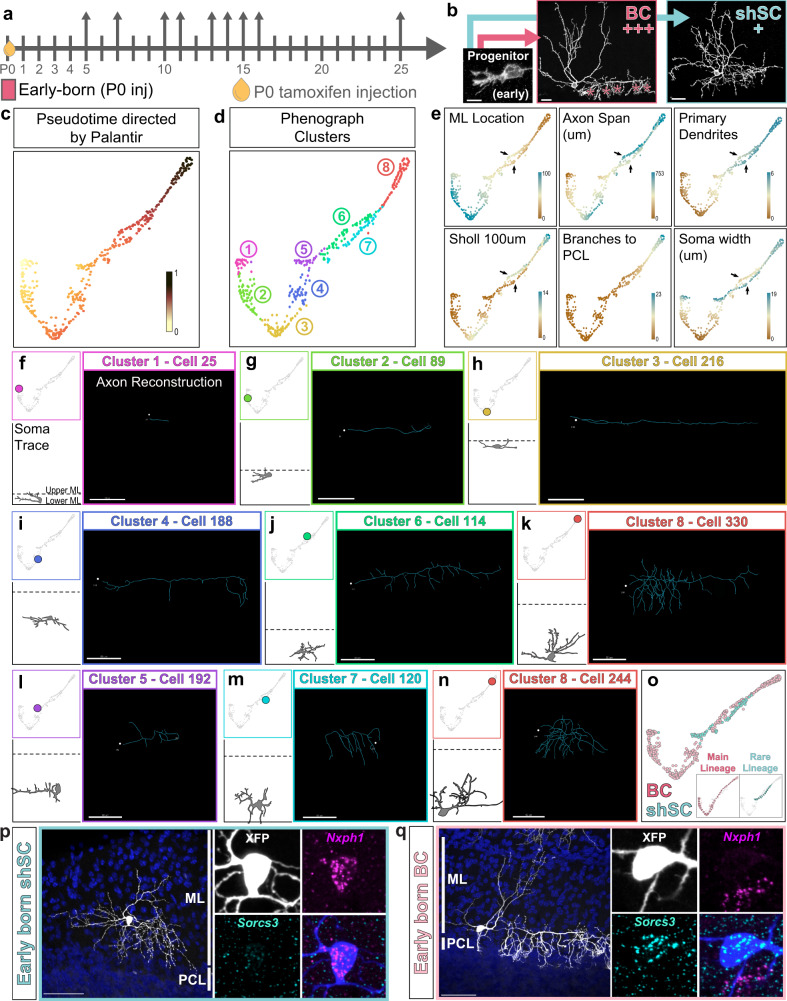
Understanding how diverse neurons are assembled into circuits requires a framework for describing cell types and their developmental trajectories. Here we combine genetic fate-mapping, pseudotemporal profiling of morphogenesis, and dual morphology and RNA labeling to resolve the diversification of mouse cerebellar inhibitory interneurons. Molecular layer interneurons (MLIs) derive from a common progenitor population but comprise diverse dendritic-, somatic-, and axon initial segment-targeting interneurons. Using quantitative morphology from 79 mature MLIs, we identify two discrete morphological types and presence of extensive within-class heterogeneity. Pseudotime trajectory inference using 732 developmental morphologies indicate the emergence of distinct MLI types during migration, before reaching their final positions. By comparing MLI identities from morphological and transcriptomic signatures, we demonstrate the dissociation between these modalities and that subtype divergence can be resolved from axonal morphogenesis prior to marker gene expression. Our study illustrates the utility of applying single-cell methods to quantify morphology for defining neuronal diversification.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


Similar articles
-
Revisiting the development of cerebellar inhibitory interneurons in the light of single-cell genetic analyses.Histochem Cell Biol. 2024 Jan;161(1):5-27. doi: 10.1007/s00418-023-02251-z. Epub 2023 Nov 8. Histochem Cell Biol. 2024. PMID: 37940705 Free PMC article. Review.
-
Concerted Interneuron Activity in the Cerebellar Molecular Layer During Rhythmic Oromotor Behaviors.J Neurosci. 2017 Nov 22;37(47):11455-11468. doi: 10.1523/JNEUROSCI.1091-17.2017. Epub 2017 Oct 24. J Neurosci. 2017. PMID: 29066561 Free PMC article.
-
Dendritic Inhibition by Shh Signaling-Dependent Stellate Cell Pool Is Critical for Motor Learning.J Neurosci. 2022 Jun 29;42(26):5130-5143. doi: 10.1523/JNEUROSCI.2073-21.2022. Epub 2022 May 19. J Neurosci. 2022. PMID: 35589396 Free PMC article.
-
Movement Rate Is Encoded and Influenced by Widespread, Coherent Activity of Cerebellar Molecular Layer Interneurons.J Neurosci. 2017 May 3;37(18):4751-4765. doi: 10.1523/JNEUROSCI.0534-17.2017. Epub 2017 Apr 7. J Neurosci. 2017. PMID: 28389475 Free PMC article.
-
Molecular layer interneurons of the cerebellum: developmental and morphological aspects.Cerebellum. 2015 Oct;14(5):534-56. doi: 10.1007/s12311-015-0648-x. Cerebellum. 2015. PMID: 25599913 Review.
Cited by
-
Distinct functional domains of Dystroglycan regulate inhibitory synapse formation and maintenance in cerebellar Purkinje cells.bioRxiv [Preprint]. 2024 Aug 30:2024.08.29.610348. doi: 10.1101/2024.08.29.610348. bioRxiv. 2024. Update in: Commun Biol. 2025 Jun 5;8(1):878. doi: 10.1038/s42003-025-08323-1. PMID: 39257744 Free PMC article. Updated. Preprint.
-
The Role of Protocadherin γ in Adult Sensory Neurons and Skin Reinnervation.J Neurosci. 2023 Dec 6;43(49):8348-8366. doi: 10.1523/JNEUROSCI.1940-22.2023. J Neurosci. 2023. PMID: 37821230 Free PMC article.
-
Perivascular neurons instruct 3D vascular lattice formation via neurovascular contact.Cell. 2024 May 23;187(11):2767-2784.e23. doi: 10.1016/j.cell.2024.04.010. Epub 2024 May 10. Cell. 2024. PMID: 38733989 Free PMC article.
-
Revisiting the development of cerebellar inhibitory interneurons in the light of single-cell genetic analyses.Histochem Cell Biol. 2024 Jan;161(1):5-27. doi: 10.1007/s00418-023-02251-z. Epub 2023 Nov 8. Histochem Cell Biol. 2024. PMID: 37940705 Free PMC article. Review.
-
Cerebellar basket cell filtering of Purkinje cell responses elicited by low frequency parallel fibre transmission.Sci Rep. 2025 Jul 12;15(1):25192. doi: 10.1038/s41598-025-09964-2. Sci Rep. 2025. PMID: 40652073 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
