

Mechanisms of structural chromosomal rearrangement formation
- PMID: 35701783
- PMCID: PMC9199198
- DOI: 10.1186/s13039-022-00600-6
Mechanisms of structural chromosomal rearrangement formation
Abstract
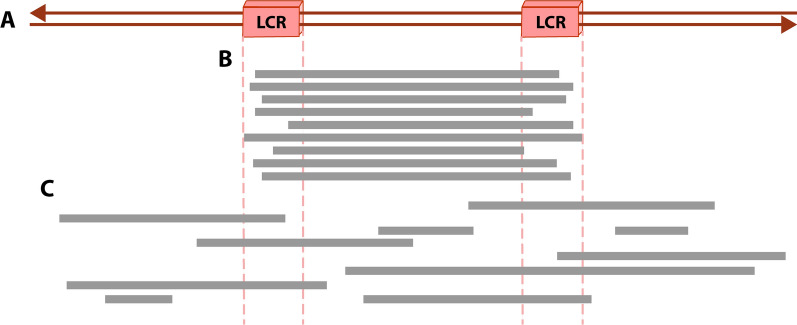
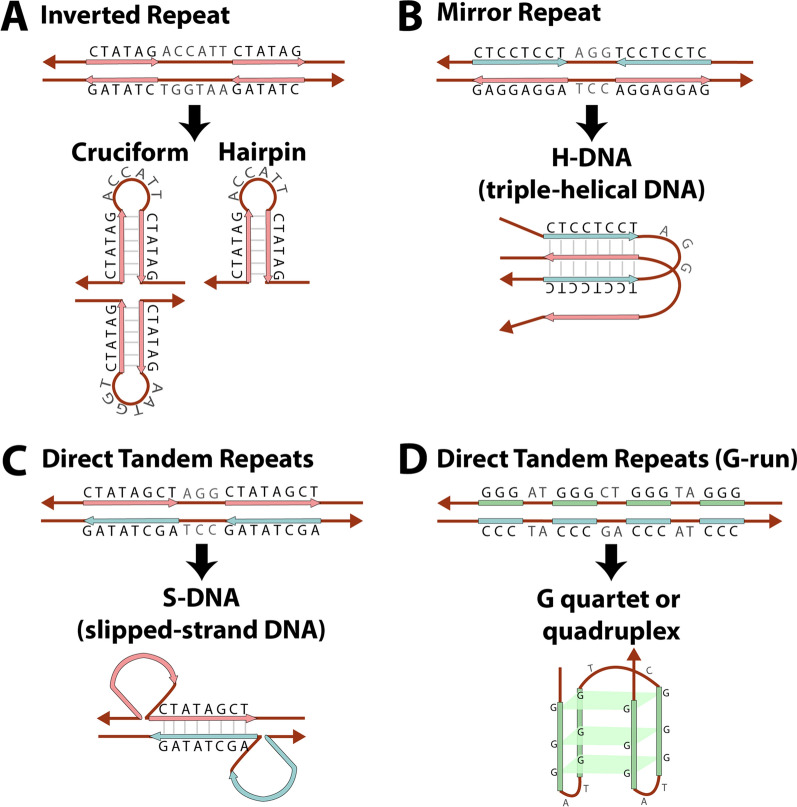
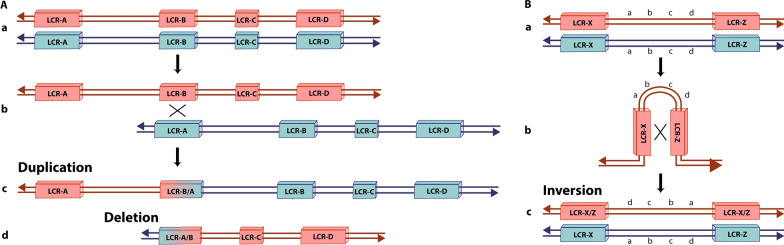
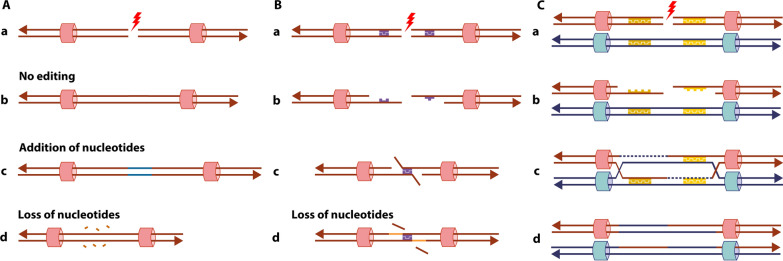
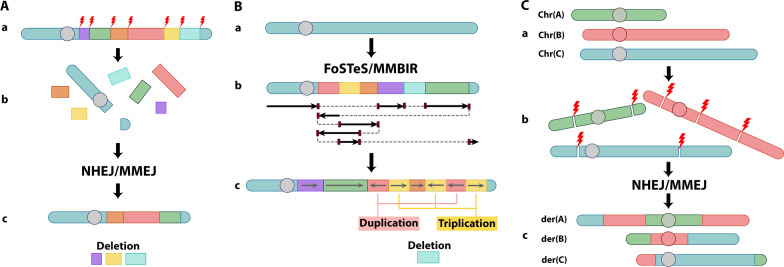
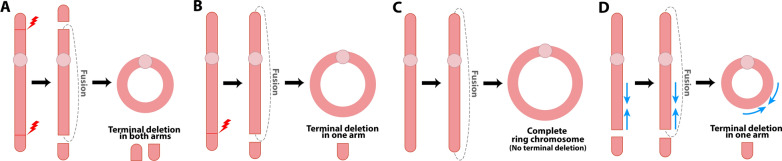
Structural chromosomal rearrangements result from different mechanisms of formation, usually related to certain genomic architectural features that may lead to genetic instability. Most of these rearrangements arise from recombination, repair, or replication mechanisms that occur after a double-strand break or the stalling/breakage of a replication fork. Here, we review the mechanisms of formation of structural rearrangements, highlighting their main features and differences. The most important mechanisms of constitutional chromosomal alterations are discussed, including Non-Allelic Homologous Recombination (NAHR), Non-Homologous End-Joining (NHEJ), Fork Stalling and Template Switching (FoSTeS), and Microhomology-Mediated Break-Induced Replication (MMBIR). Their involvement in chromoanagenesis and in the formation of complex chromosomal rearrangements, inverted duplications associated with terminal deletions, and ring chromosomes is also outlined. We reinforce the importance of high-resolution analysis to determine the DNA sequence at, and near, their breakpoints in order to infer the mechanisms of formation of structural rearrangements and to reveal how cells respond to DNA damage and repair broken ends.
Keywords: Chromoanagenesis; Fork stalling and template switching (FoSTeS); Inv dup del; Mechanisms of formation; Microhomology-mediated break-induced replication (MMBIR); Non-allelic homologous recombination (NAHR); Non-homologous end-joining (NHEJ); Ring chromosome; Structural chromosomal rearrangements.
© 2022. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures



References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
