

Hematopoietic stem cell regeneration through paracrine regulation of the Wnt5a/Prox1 signaling axis
- PMID: 35703178
- PMCID: PMC9197516
- DOI: 10.1172/JCI155914
Hematopoietic stem cell regeneration through paracrine regulation of the Wnt5a/Prox1 signaling axis
Abstract
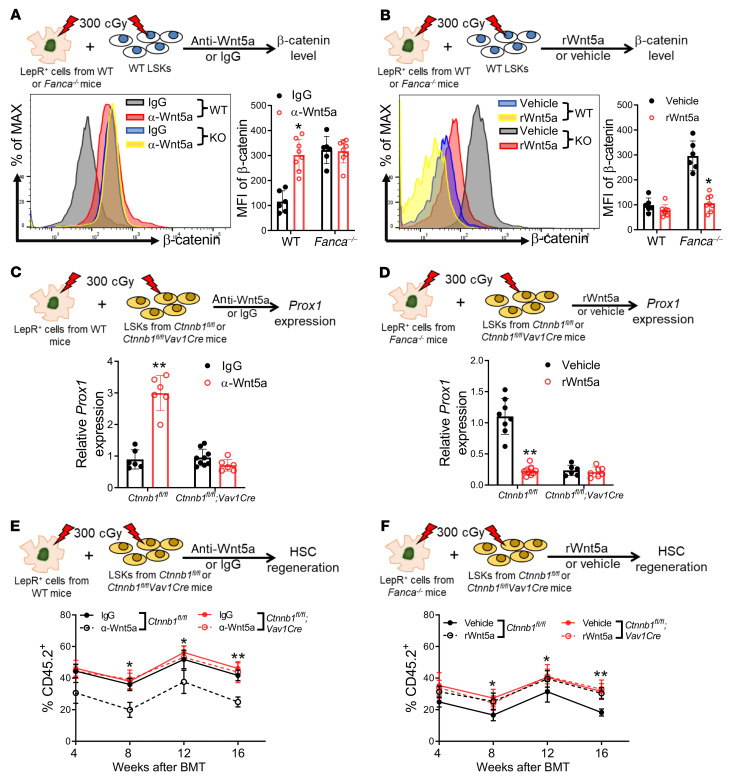
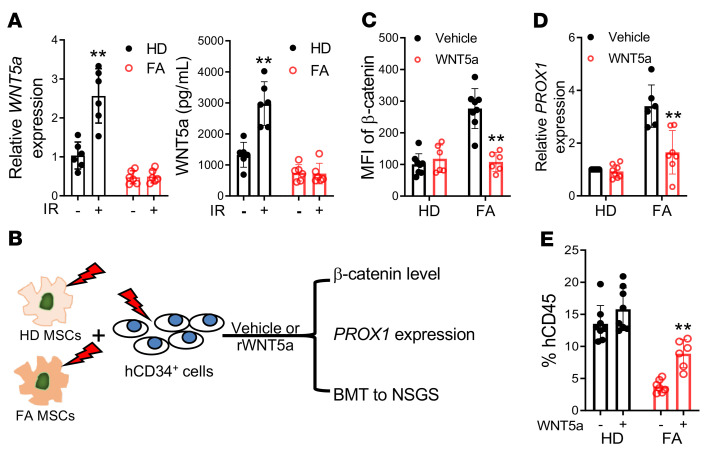
The crosstalk between the BM microenvironment (niche) and hematopoietic stem cells (HSCs) is critical for HSC regeneration. Here, we show that in mice, deletion of the Fanconi anemia (FA) genes Fanca and Fancc dampened HSC regeneration through direct effects on HSCs and indirect effects on BM niche cells. FA HSCs showed persistent upregulation of the Wnt target Prox1 in response to total body irradiation (TBI). Accordingly, lineage-specific deletion of Prox1 improved long-term repopulation of the irradiated FA HSCs. Forced expression of Prox1 in WT HSCs mimicked the defective repopulation phenotype of FA HSCs. WT mice but not FA mice showed significant induction by TBI of BM stromal Wnt5a protein. Mechanistically, FA proteins regulated stromal Wnt5a expression, possibly through modulating the Wnt5a transcription activator Pax2. Wnt5a treatment of irradiated FA mice enhanced HSC regeneration. Conversely, Wnt5a neutralization inhibited HSC regeneration after TBI. Wnt5a secreted by LepR+CXCL12+ BM stromal cells inhibited β-catenin accumulation, thereby repressing Prox1 transcription in irradiated HSCs. The detrimental effect of deregulated Wnt5a/Prox1 signaling on HSC regeneration was also observed in patients with FA and aged mice. Irradiation induced upregulation of Prox1 in the HSCs of aged mice, and deletion of Prox1 in aged HSCs improved HSC regeneration. Treatment of aged mice with Wnt5a enhanced hematopoietic repopulation. Collectively, these findings identified the paracrine Wnt5a/Prox1 signaling axis as a regulator of HSC regeneration under conditions of injury and aging.
Keywords: Cell Biology; Hematology; Hematopoietic stem cells.
Figures






Similar articles
-
p53-TP53-Induced Glycolysis Regulator Mediated Glycolytic Suppression Attenuates DNA Damage and Genomic Instability in Fanconi Anemia Hematopoietic Stem Cells.Stem Cells. 2019 Jul;37(7):937-947. doi: 10.1002/stem.3015. Epub 2019 May 3. Stem Cells. 2019. PMID: 30977208 Free PMC article.
-
Wnt5a regulates hematopoietic stem cell proliferation and repopulation through the Ryk receptor.Stem Cells. 2014 Jan;32(1):105-15. doi: 10.1002/stem.1513. Stem Cells. 2014. PMID: 23939973 Free PMC article.
-
Wnt5a inhibits canonical Wnt signaling in hematopoietic stem cells and enhances repopulation.Proc Natl Acad Sci U S A. 2007 Sep 25;104(39):15436-41. doi: 10.1073/pnas.0704747104. Epub 2007 Sep 19. Proc Natl Acad Sci U S A. 2007. PMID: 17881570 Free PMC article.
-
Wnt-5A/B Signaling in Hematopoiesis throughout Life.Cells. 2020 Jul 29;9(8):1801. doi: 10.3390/cells9081801. Cells. 2020. PMID: 32751131 Free PMC article. Review.
-
Stem Cell Genetic Therapy for Fanconi Anemia - A New Hope.Curr Gene Ther. 2017;16(5):309-320. doi: 10.2174/1566523217666170109111958. Curr Gene Ther. 2017. PMID: 28067166 Review.
Cited by
-
M1-BMDMs with Wnt5a deletion attenuate liver fibrosis by suppression of Wnt5a/Frizzled 2 axis in hepatic progenitors.Cell Biosci. 2025 Aug 30;15(1):125. doi: 10.1186/s13578-025-01467-x. Cell Biosci. 2025. PMID: 40885967 Free PMC article.
-
HES1 is required for mouse fetal hematopoiesis.Stem Cell Res Ther. 2024 Jul 29;15(1):235. doi: 10.1186/s13287-024-03836-8. Stem Cell Res Ther. 2024. PMID: 39075526 Free PMC article.
-
Lymphatic vasculature in the central nervous system.Front Cell Dev Biol. 2023 Apr 7;11:1150775. doi: 10.3389/fcell.2023.1150775. eCollection 2023. Front Cell Dev Biol. 2023. PMID: 37091974 Free PMC article. Review.
-
PGE2 Produced by Exogenous MSCs Promotes Immunoregulation in ARDS Induced by Highly Pathogenic Influenza A through Activation of the Wnt-β-Catenin Signaling Pathway.Int J Mol Sci. 2023 Apr 14;24(8):7299. doi: 10.3390/ijms24087299. Int J Mol Sci. 2023. PMID: 37108459 Free PMC article. Review.
-
The quest for the holy grail: overcoming challenges in expanding human hematopoietic stem cells for clinical use.Stem Cell Investig. 2023 Jul 11;10:15. doi: 10.21037/sci-2023-016. eCollection 2023. Stem Cell Investig. 2023. PMID: 37457748 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
