

Calreticulin mutant myeloproliferative neoplasms induce MHC-I skewing, which can be overcome by an optimized peptide cancer vaccine
- PMID: 35704596
- PMCID: PMC11182673
- DOI: 10.1126/scitranslmed.aba4380
Calreticulin mutant myeloproliferative neoplasms induce MHC-I skewing, which can be overcome by an optimized peptide cancer vaccine
Abstract
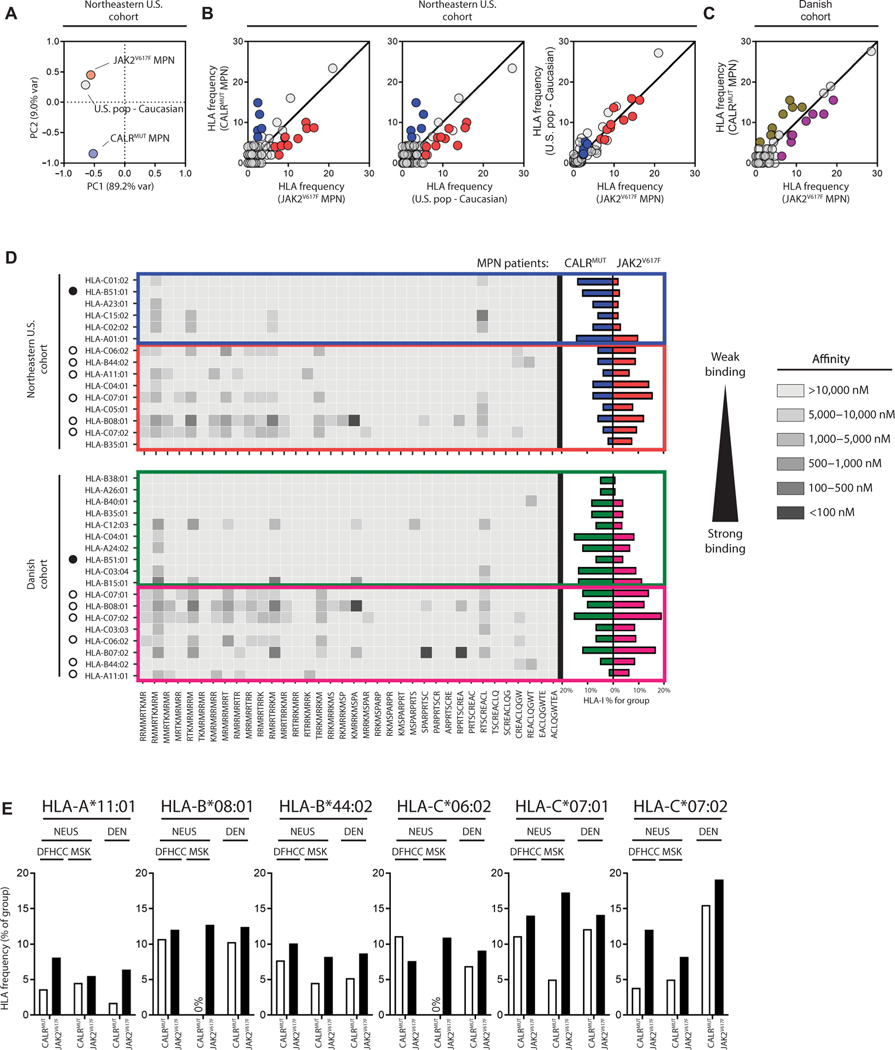
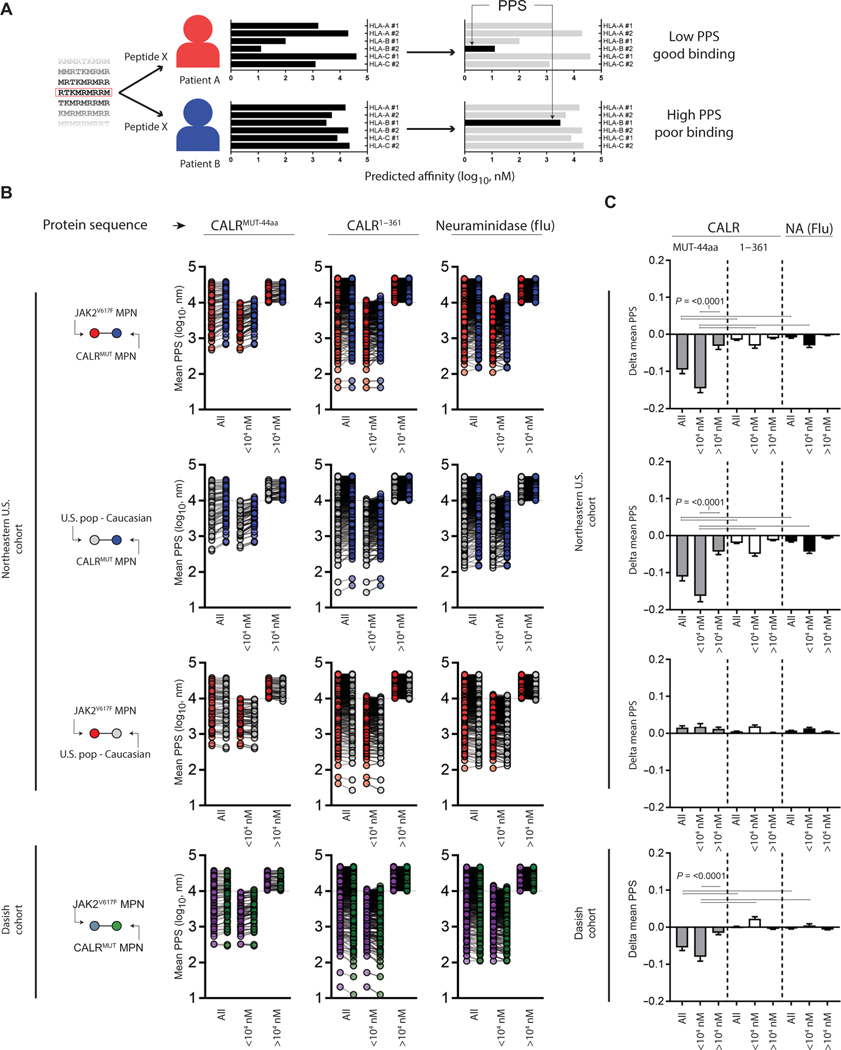
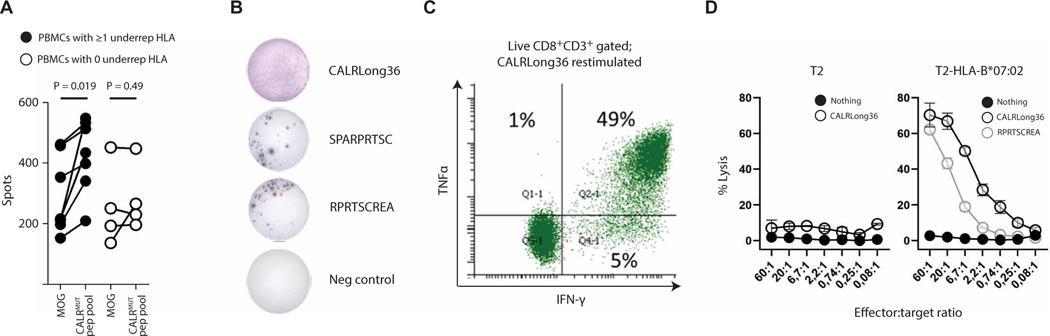
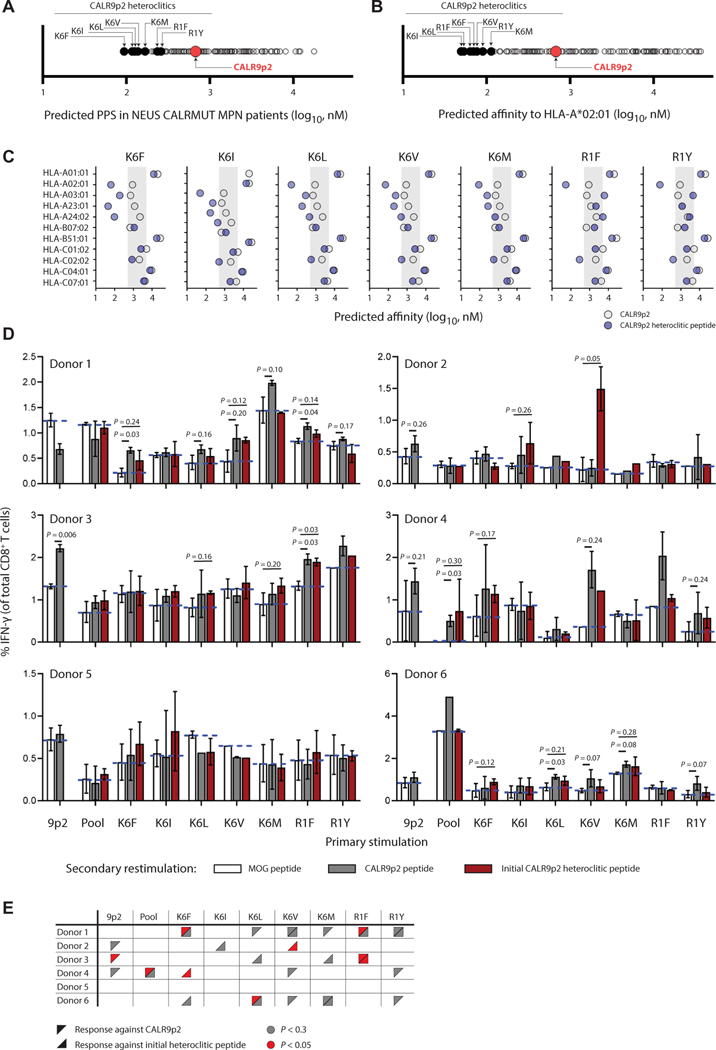
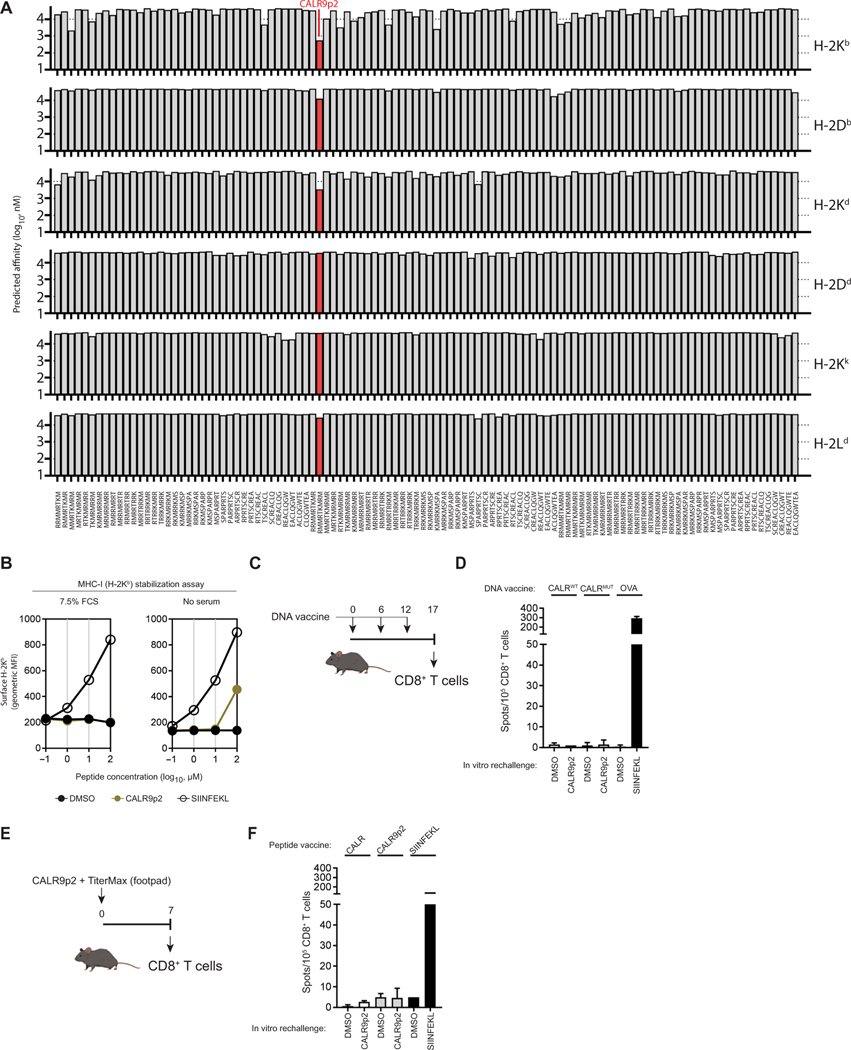
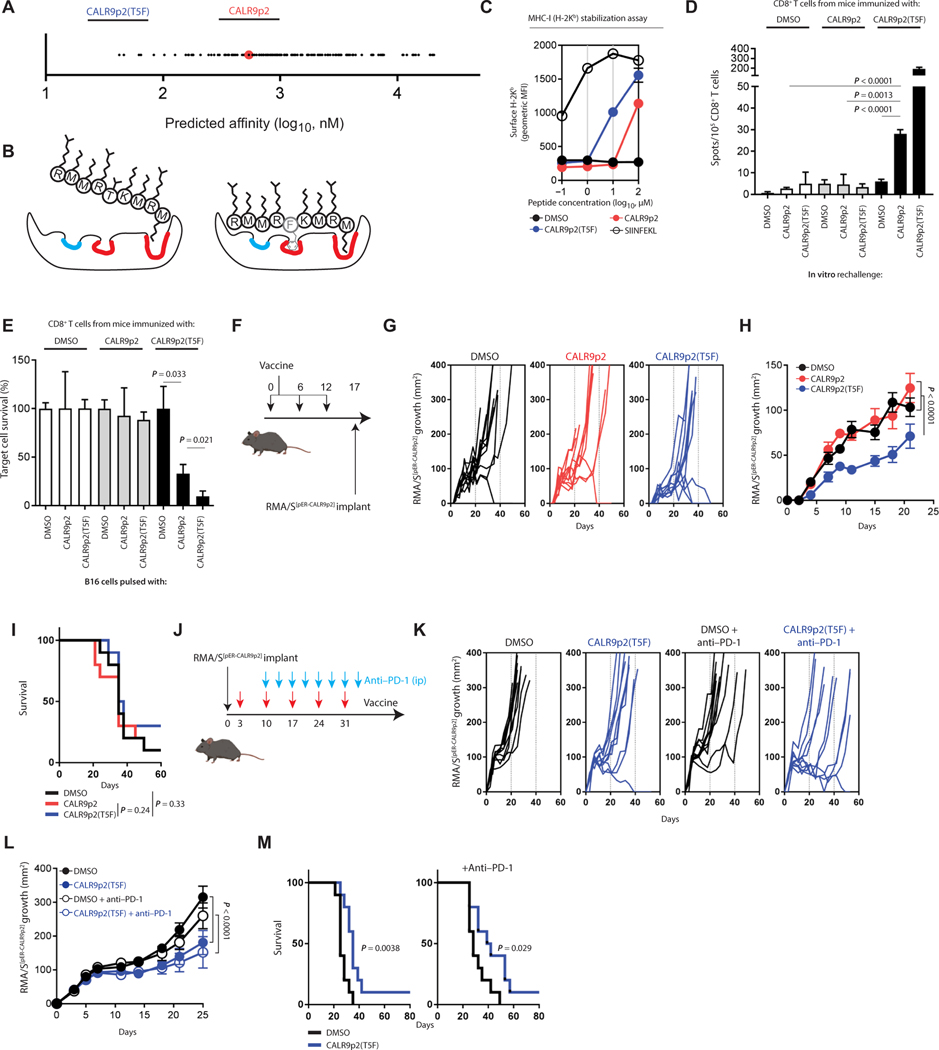
The majority of JAK2V617F-negative myeloproliferative neoplasms (MPNs) have disease-initiating frameshift mutations in calreticulin (CALR), resulting in a common carboxyl-terminal mutant fragment (CALRMUT), representing an attractive source of neoantigens for cancer vaccines. However, studies have shown that CALRMUT-specific T cells are rare in patients with CALRMUT MPN for unknown reasons. We examined class I major histocompatibility complex (MHC-I) allele frequencies in patients with CALRMUT MPN from two independent cohorts. We observed that MHC-I alleles that present CALRMUT neoepitopes with high affinity are underrepresented in patients with CALRMUT MPN. We speculated that this was due to an increased chance of immune-mediated tumor rejection by individuals expressing one of these MHC-I alleles such that the disease never clinically manifested. As a consequence of this MHC-I allele restriction, we reasoned that patients with CALRMUT MPN would not efficiently respond to a CALRMUT fragment cancer vaccine but would when immunized with a modified CALRMUT heteroclitic peptide vaccine approach. We found that heteroclitic CALRMUT peptides specifically designed for the MHC-I alleles of patients with CALRMUT MPN efficiently elicited a CALRMUT cross-reactive CD8+ T cell response in human peripheral blood samples but not to the matched weakly immunogenic CALRMUT native peptides. We corroborated this effect in vivo in mice and observed that C57BL/6J mice can mount a CD8+ T cell response to the CALRMUT fragment upon immunization with a CALRMUT heteroclitic, but not native, peptide. Together, our data emphasize the therapeutic potential of heteroclitic peptide-based cancer vaccines in patients with CALRMUT MPN.
Figures
References
-
- Campbell PJ, Green AR, The myeloproliferative disorders. N. Engl. J. Med. 355, 2452–2466 (2006). - PubMed
-
- Rampal R, Al-Shahrour F, Abdel-Wahab O, Patel JP, Brunel JP, Mermel CH, Bass AJ, Pretz J, Ahn J, Hricik T, Kilpivaara O, Wadleigh M, Busque L, Gilliland DG, Golub TR, Ebert BL, Levine RL, Integrated genomic analysis illustrates the central role of JAK-STAT pathway activation in myeloproliferative neoplasm pathogenesis. Blood 123, e123–e133 (2014). - PMC - PubMed
-
- Nangalia J, Massie CE, Baxter EJ, Nice FL, Gundem G, Wedge DC, Avezov E, Li J, Kollmann K, Kent DG, Aziz A, Godfrey AL, Hinton J, Martincorena I, Van Loo P, Jones AV, Guglielmelli P, Tarpey P, Harding HP, Fitzpatrick JD, Goudie CT, Ortmann CA, Loughran SJ, Raine K, Jones DR, Butler AP, Teague JW, O’Meara S, McLaren S, Bianchi M, Silber Y, Dimitropoulou D, Bloxham D, Mudie L, Maddison M, Robinson B, Keohane C, Maclean C, Hill K, Orchard K, Tauro S, Du MQ, Greaves M, Bowen BJP Huntly CN Harrison NC Cross P, Ron D, Vannucchi AM, Papaemmanuil, Campbell PJ, Green AR, Somatic CALR mutations in myeloproliferative neoplasms with nonmutated JAK2. N. Engl. J. Med. 369, 2391–2405 (2013). - PMC - PubMed
-
- Klampfl T, Gisslinger H, Harutyunyan AS, Nivarthi H, Rumi E, Milosevic JD, Them NC, Berg T, Gisslinger B, Pietra D, Chen D, Vladimer GI, Bagienski K, Milanesi C, Casetti IC, Sant’Antonio E, Ferretti V, Elena C, Schischlik F, Cleary C, Six M, Schalling M, Schonegger A, Bock C, Malcovati L, Pascutto C, Superti-Furga G, Cazzola M, Kralovics R, Somatic mutations of calreticulin in myeloproliferative neoplasms. N. Engl. J. Med. 369, 2379–2390 (2013). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
