

The balance between protective and pathogenic immune responses to pneumonia in the neonatal lung is enforced by gut microbiota
- PMID: 35704600
- PMCID: PMC10032669
- DOI: 10.1126/scitranslmed.abl3981
The balance between protective and pathogenic immune responses to pneumonia in the neonatal lung is enforced by gut microbiota
Abstract
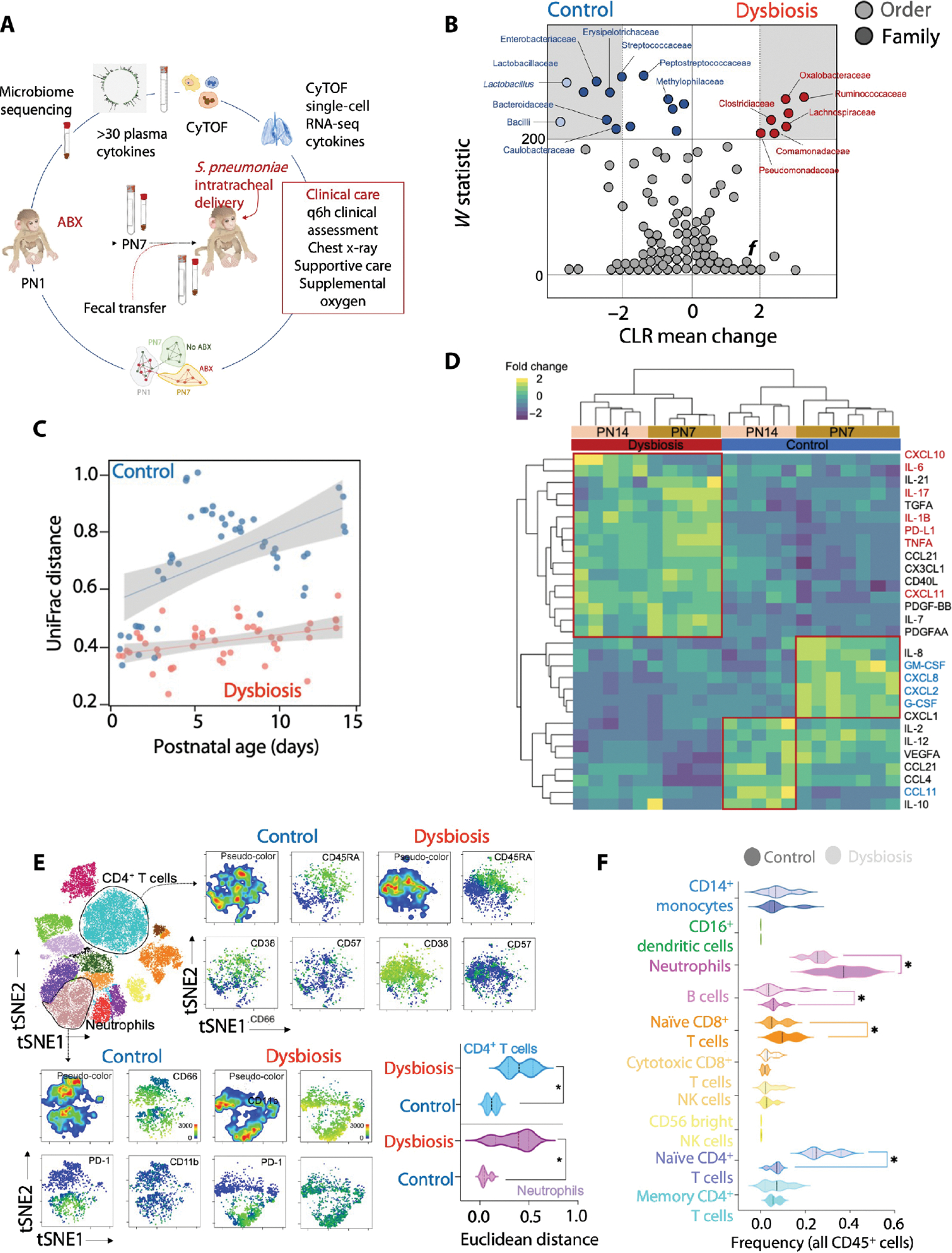
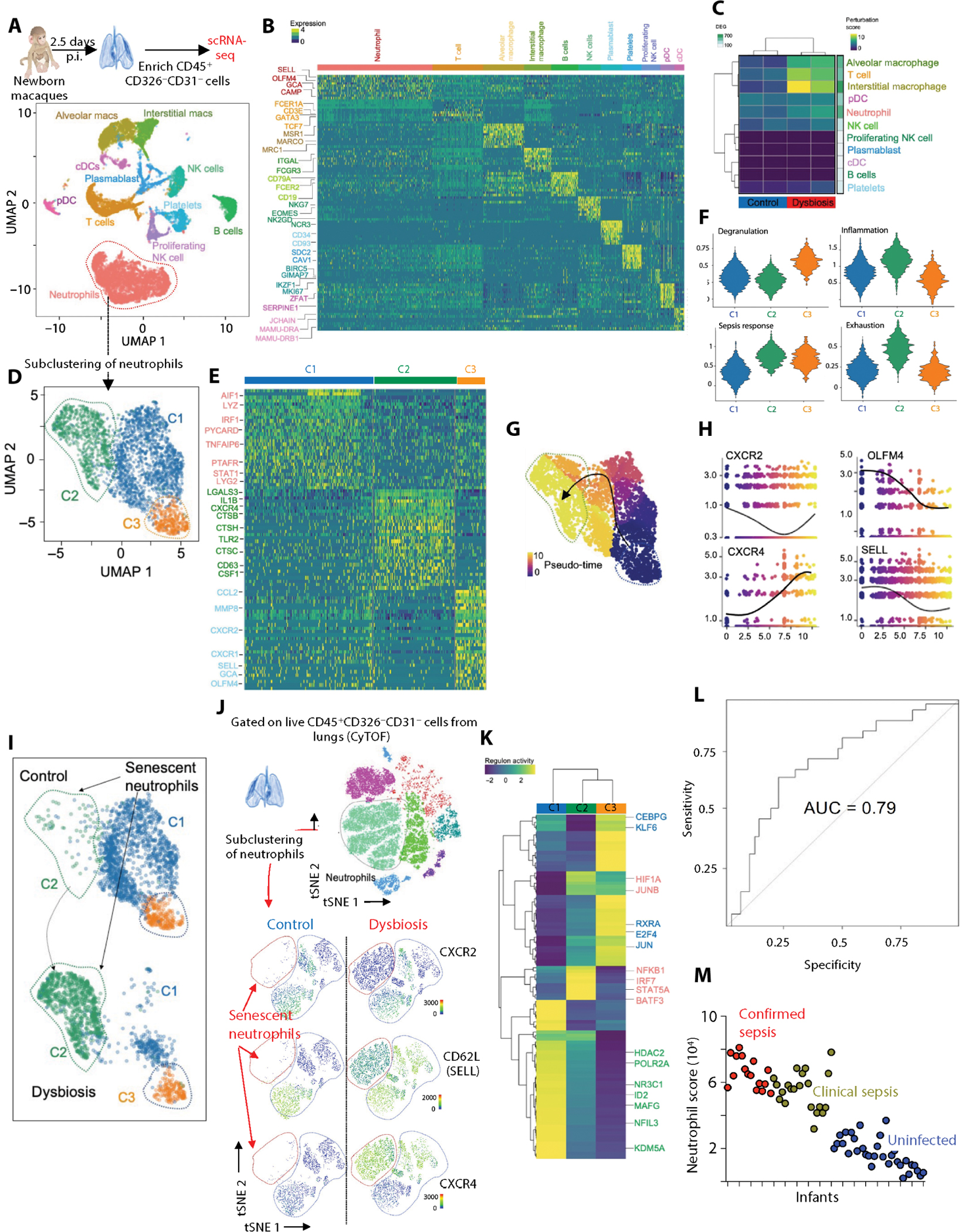
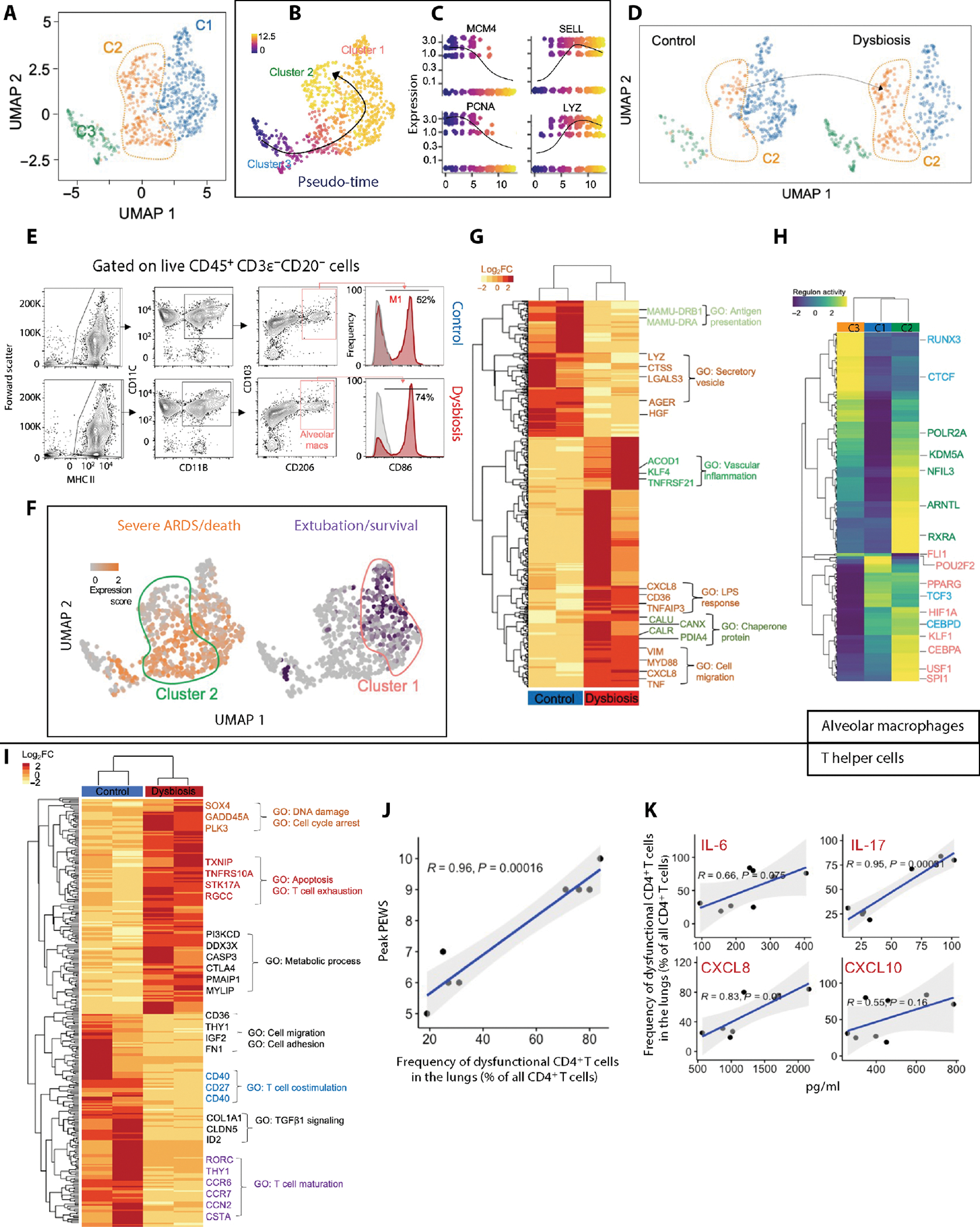
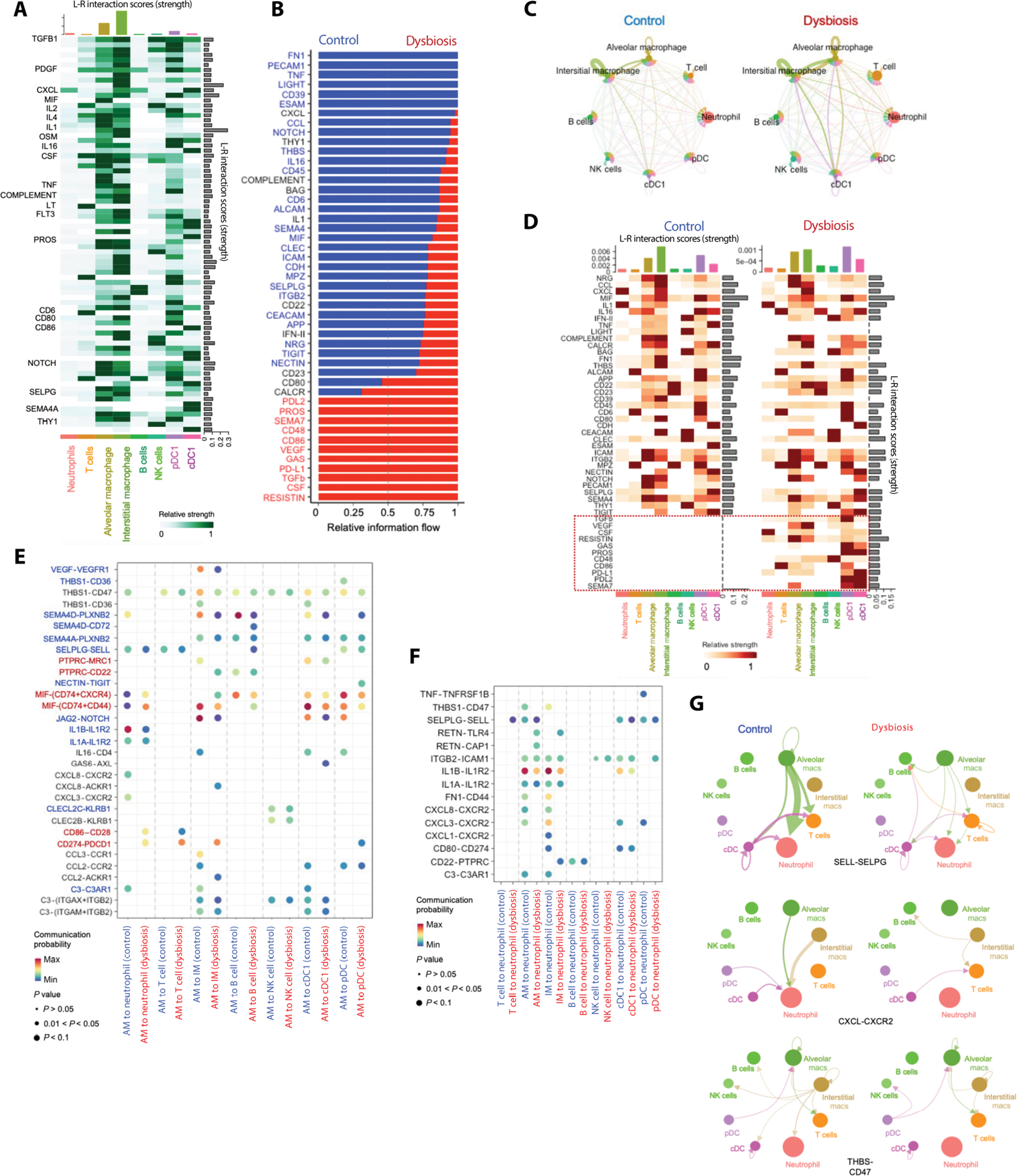
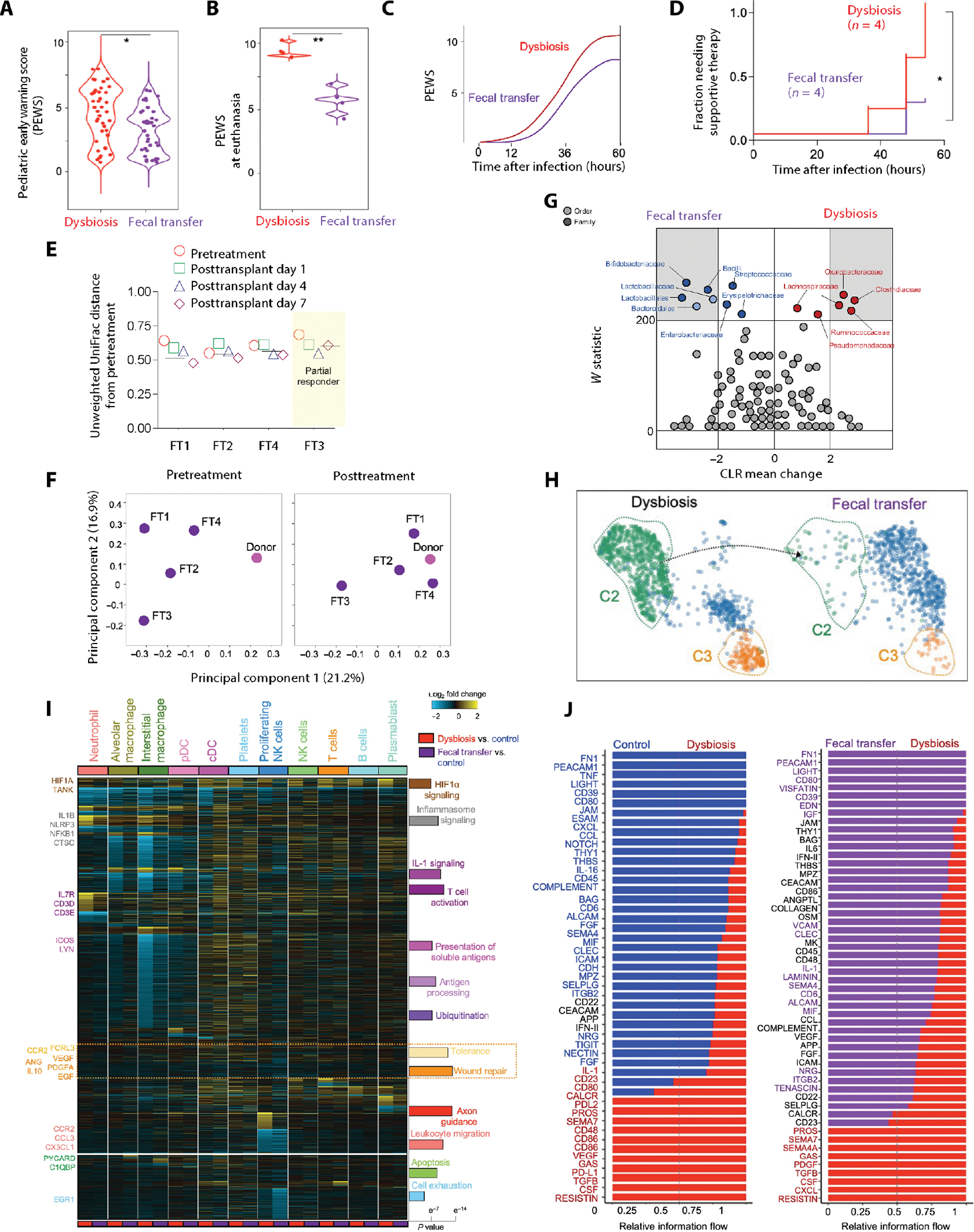
Although modern clinical practices such as cesarean sections and perinatal antibiotics have improved infant survival, treatment with broad-spectrum antibiotics alters intestinal microbiota and causes dysbiosis. Infants exposed to perinatal antibiotics have an increased likelihood of life-threatening infections, including pneumonia. Here, we investigated how the gut microbiota sculpt pulmonary immune responses, promoting recovery and resolution of infection in newborn rhesus macaques. Early-life antibiotic exposure interrupted the maturation of intestinal commensal bacteria and disrupted the developmental trajectory of the pulmonary immune system, as assessed by single-cell proteomic and transcriptomic analyses. Early-life antibiotic exposure rendered newborn macaques more susceptible to bacterial pneumonia, concurrent with increases in neutrophil senescence and hyperinflammation, broad inflammatory cytokine signaling, and macrophage dysfunction. This pathogenic reprogramming of pulmonary immunity was further reflected by a hyperinflammatory signature in all pulmonary immune cell subsets coupled with a global loss of tissue-protective, homeostatic pathways in the lungs of dysbiotic newborns. Fecal microbiota transfer was associated with partial correction of the broad immune maladaptations and protection against severe pneumonia. These data demonstrate the importance of intestinal microbiota in programming pulmonary immunity and support the idea that gut microbiota promote the balance between pathways driving tissue repair and inflammatory responses associated with clinical recovery from infection in infants. Our results highlight a potential role for microbial transfer for immune support in these at-risk infants.
Conflict of interest statement
Figures

Similar articles
-
Dysbiosis of Gut Microbiota and Intestinal Barrier Dysfunction in Pigs with Pulmonary Inflammation Induced by Mycoplasma hyorhinis Infection.mSystems. 2022 Aug 30;7(4):e0028222. doi: 10.1128/msystems.00282-22. Epub 2022 Jun 14. mSystems. 2022. PMID: 35699454 Free PMC article.
-
Perinatal Antibiotic Exposure Affects the Transmission between Maternal and Neonatal Microbiota and Is Associated with Early-Onset Sepsis.mSphere. 2020 Feb 19;5(1):e00984-19. doi: 10.1128/mSphere.00984-19. mSphere. 2020. PMID: 32075882 Free PMC article.
-
An Antibiotic-Impacted Microbiota Compromises the Development of Colonic Regulatory T Cells and Predisposes to Dysregulated Immune Responses.mBio. 2021 Feb 2;12(1):e03335-20. doi: 10.1128/mBio.03335-20. mBio. 2021. PMID: 33531385 Free PMC article.
-
The developing gut microbiota and its consequences for health.J Dev Orig Health Dis. 2018 Dec;9(6):590-597. doi: 10.1017/S2040174418000119. Epub 2018 Mar 22. J Dev Orig Health Dis. 2018. PMID: 29562949 Review.
-
The impact of maternal exposure to antibiotics on the development of child gut microbiome.Immunol Med. 2022 Jun;45(2):63-68. doi: 10.1080/25785826.2021.1963189. Epub 2021 Aug 15. Immunol Med. 2022. PMID: 34392799 Review.
Cited by
-
Prevention and alleviation of allergic rhinitis by oral administration of Lacticaseibacillus paracasei GOLDGUT-Lpc969.Front Immunol. 2024 Sep 30;15:1444778. doi: 10.3389/fimmu.2024.1444778. eCollection 2024. Front Immunol. 2024. PMID: 39403377 Free PMC article.
-
The gut microbiome as a potential source of non-invasive biomarkers of chronic obstructive pulmonary disease.Front Microbiol. 2023 Jul 25;14:1173614. doi: 10.3389/fmicb.2023.1173614. eCollection 2023. Front Microbiol. 2023. PMID: 37555072 Free PMC article.
-
Less is more: Antibiotics at the beginning of life.Nat Commun. 2023 Apr 27;14(1):2423. doi: 10.1038/s41467-023-38156-7. Nat Commun. 2023. PMID: 37105958 Free PMC article. Review.
-
Extra-pulmonary control of respiratory defense.Cell Immunol. 2024 Jul-Aug;401-402:104841. doi: 10.1016/j.cellimm.2024.104841. Epub 2024 Jun 7. Cell Immunol. 2024. PMID: 38878619 Free PMC article. Review.
-
Making friends: active selection of symbionts and rejection of pathogens by the neonatal immune system.Front Immunol. 2023 Nov 20;14:1287518. doi: 10.3389/fimmu.2023.1287518. eCollection 2023. Front Immunol. 2023. PMID: 38054002 Free PMC article.
References
-
- Elinav E, Strowig T, Henao-Mejia J, Flavell RA, Regulation of the antimicrobial response by NLR proteins. Immunity 34, 665–679 (2011). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 HL142708/HL/NHLBI NIH HHS/United States
- R43 EB029863/EB/NIBIB NIH HHS/United States
- R01 HL149366/HL/NHLBI NIH HHS/United States
- R21 HL142485/HL/NHLBI NIH HHS/United States
- U24 HL148865/HL/NHLBI NIH HHS/United States
- P51 OD011107/OD/NIH HHS/United States
- P30 DK078392/DK/NIDDK NIH HHS/United States
- R01 AI150554/AI/NIAID NIH HHS/United States
- R01 AI143554/AI/NIAID NIH HHS/United States
- R21 AI152100/AI/NIAID NIH HHS/United States
- P30 DK090971/DK/NIDDK NIH HHS/United States
- R01 AI138553/AI/NIAID NIH HHS/United States
- R01 CA226802/CA/NCI NIH HHS/United States
- P30 AR047363/AR/NIAMS NIH HHS/United States
- U42 OD027094/OD/NIH HHS/United States
- R01 GM128452/GM/NIGMS NIH HHS/United States
- R01 HD089939/HD/NICHD NIH HHS/United States
- U01 HL134745/HL/NHLBI NIH HHS/United States
- K08 HL140178/HL/NHLBI NIH HHS/United States
- K08 HD084686/HD/NICHD NIH HHS/United States
- R41 AI157626/AI/NIAID NIH HHS/United States
- K12 HD028827/HD/NICHD NIH HHS/United States
- U01 ES029234/ES/NIEHS NIH HHS/United States
- R01 HL155611/HL/NHLBI NIH HHS/United States
- U01 HL122642/HL/NHLBI NIH HHS/United States
- R01 AG053498/AG/NIA NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
