

Realization of the T Lineage Program Involves GATA-3 Induction of Bcl11b and Repression of Cdkn2b Expression
- PMID: 35705252
- PMCID: PMC9248976
- DOI: 10.4049/jimmunol.2100366
Realization of the T Lineage Program Involves GATA-3 Induction of Bcl11b and Repression of Cdkn2b Expression
Abstract
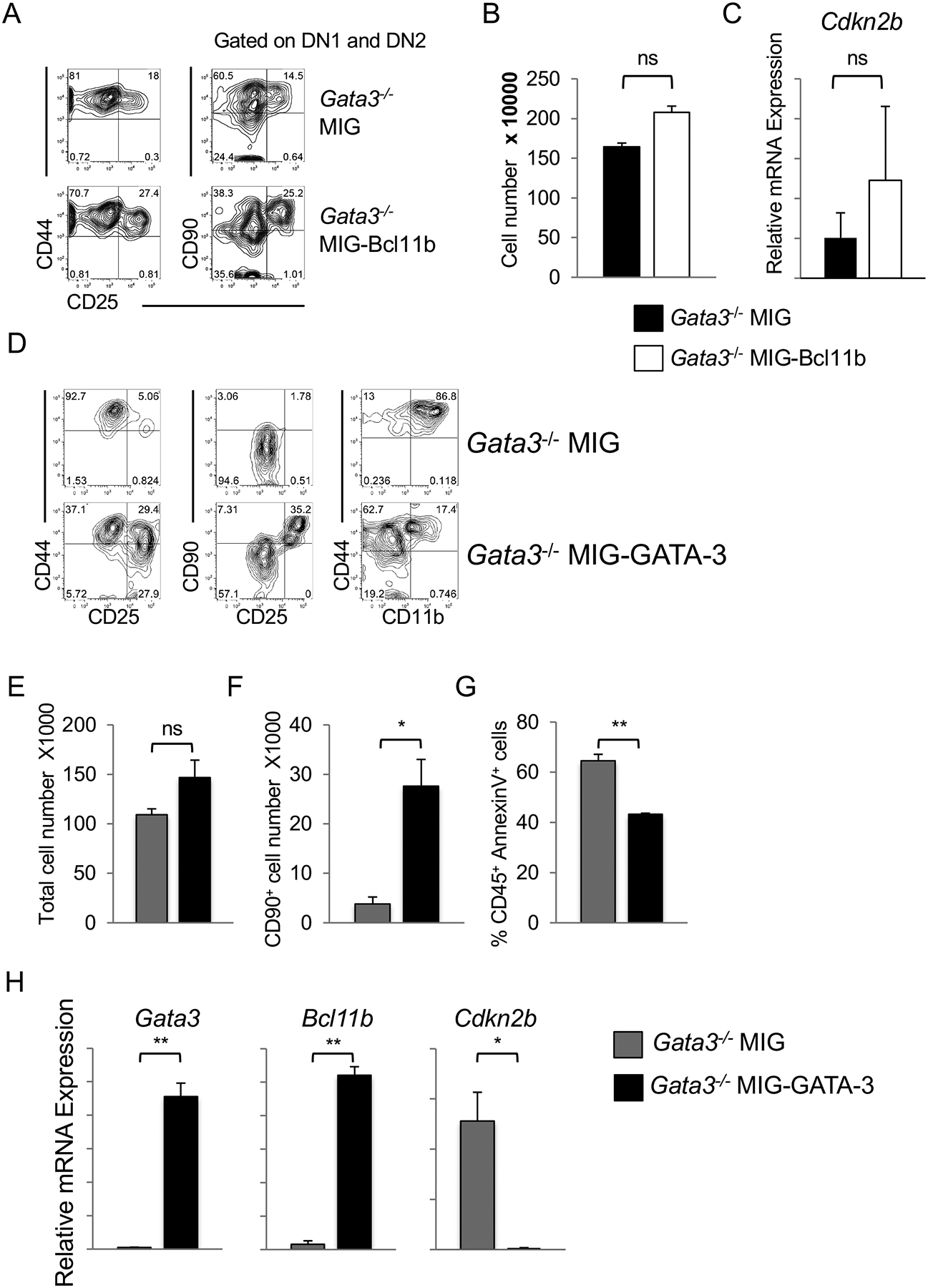
The zinc-finger transcription factor GATA-3 plays a crucial role during early T cell development and also dictates later T cell differentiation outcomes. However, its role and collaboration with the Notch signaling pathway in the induction of T lineage specification and commitment have not been fully elucidated. We show that GATA-3 deficiency in mouse hematopoietic progenitors results in an early block in T cell development despite the presence of Notch signals, with a failure to upregulate Bcl11b expression, leading to a diversion along a myeloid, but not a B cell, lineage fate. GATA-3 deficiency in the presence of Notch signaling results in the apoptosis of early T lineage cells, as seen with inhibition of CDK4/6 (cyclin-dependent kinases 4 and 6) function, and dysregulated cyclin-dependent kinase inhibitor 2b (Cdkn2b) expression. We also show that GATA-3 induces Bcl11b, and together with Bcl11b represses Cdkn2b expression; however, loss of Cdkn2b failed to rescue the developmental block of GATA-3-deficient T cell progenitor. Our findings provide a signaling and transcriptional network by which the T lineage program in response to Notch signals is realized.
Copyright © 2022 by The American Association of Immunologists, Inc.
Figures







References
-
- Petrie HT, and Zuniga-Pflucker JC. 2007. Zoned out: functional mapping of stromal signaling microenvironments in the thymus. Annu Rev Immunol 25: 649–679. - PubMed
-
- Bhandoola A, von Boehmer H, Petrie HT, and Zuniga-Pflucker JC. 2007. Commitment and developmental potential of extrathymic and intrathymic T cell precursors: plenty to choose from. Immunity 26: 678–689. - PubMed
-
- Sambandam A, Maillard I, Zediak VP, Xu L, Gerstein RM, Aster JC, Pear WS, and Bhandoola A. 2005. Notch signaling controls the generation and differentiation of early T lineage progenitors. Nat Immunol 6: 663–670. - PubMed
-
- Heinzel K, Benz C, Martins VC, Haidl ID, and Bleul CC. 2007. Bone marrow-derived hemopoietic precursors commit to the T cell lineage only after arrival in the thymic microenvironment. J Immunol 178: 858–868. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
