

Ly6D+Siglec-H+ precursors contribute to conventional dendritic cells via a Zbtb46+Ly6D+ intermediary stage
- PMID: 35705536
- PMCID: PMC9200809
- DOI: 10.1038/s41467-022-31054-4
Ly6D+Siglec-H+ precursors contribute to conventional dendritic cells via a Zbtb46+Ly6D+ intermediary stage
Abstract
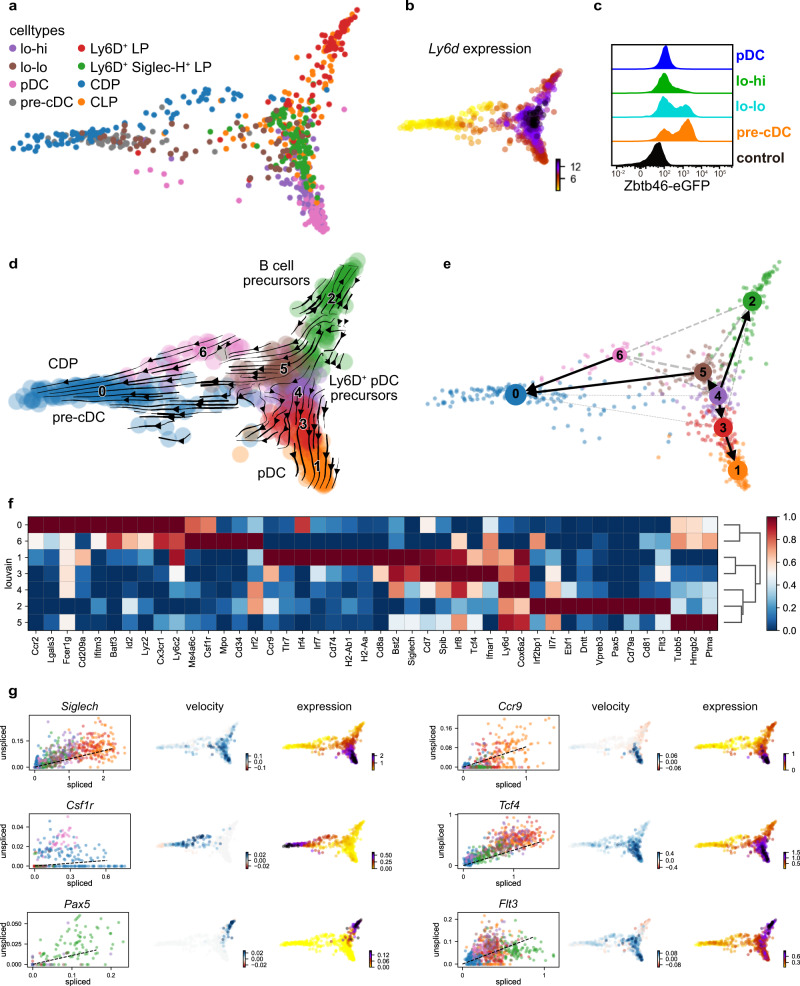
Plasmacytoid and conventional dendritic cells (pDC and cDC) are generated from progenitor cells in the bone marrow and commitment to pDCs or cDC subtypes may occur in earlier and later progenitor stages. Cells within the CD11c+MHCII-/loSiglec-H+CCR9lo DC precursor fraction of the mouse bone marrow generate both pDCs and cDCs. Here we investigate the heterogeneity and commitment of subsets in this compartment by single-cell transcriptomics and high-dimensional flow cytometry combined with cell fate analysis: Within the CD11c+MHCII-/loSiglec-H+CCR9lo DC precursor pool cells expressing high levels of Ly6D and lacking expression of transcription factor Zbtb46 contain CCR9loB220hi immediate pDC precursors and CCR9loB220lo (lo-lo) cells which still generate pDCs and cDCs in vitro and in vivo under steady state conditions. cDC-primed cells within the Ly6DhiZbtb46- lo-lo precursors rapidly upregulate Zbtb46 and pass through a Zbtb46+Ly6D+ intermediate stage before acquiring cDC phenotype after cell division. Type I IFN stimulation limits cDC and promotes pDC output from this precursor fraction by arresting cDC-primed cells in the Zbtb46+Ly6D+ stage preventing their expansion and differentiation into cDCs. Modulation of pDC versus cDC output from precursors by external factors may allow for adaptation of DC subset composition at later differentiation stages.
© 2022. The Author(s).
Conflict of interest statement
F.J.T. reports receiving consulting fees from Roche Diagnostics GmbH, Immunai Inc., Singularity Bio B.V., CytoReason Ltd, and Omniscope Ltd Inc., and has an ownership interest in Cellarity, Inc. and Dermagnostix GmbH. VB is a full-time employee of Cellarity Inc. and reports ownership interest in Cellarity Inc.; the present work was carried out at Helmholtz Munich. The other authors declare no competing interests.
Figures





References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
