

Placental sex-dependent spermine synthesis regulates trophoblast gene expression through acetyl-coA metabolism and histone acetylation
- PMID: 35705689
- PMCID: PMC9200719
- DOI: 10.1038/s42003-022-03530-6
Placental sex-dependent spermine synthesis regulates trophoblast gene expression through acetyl-coA metabolism and histone acetylation
Abstract
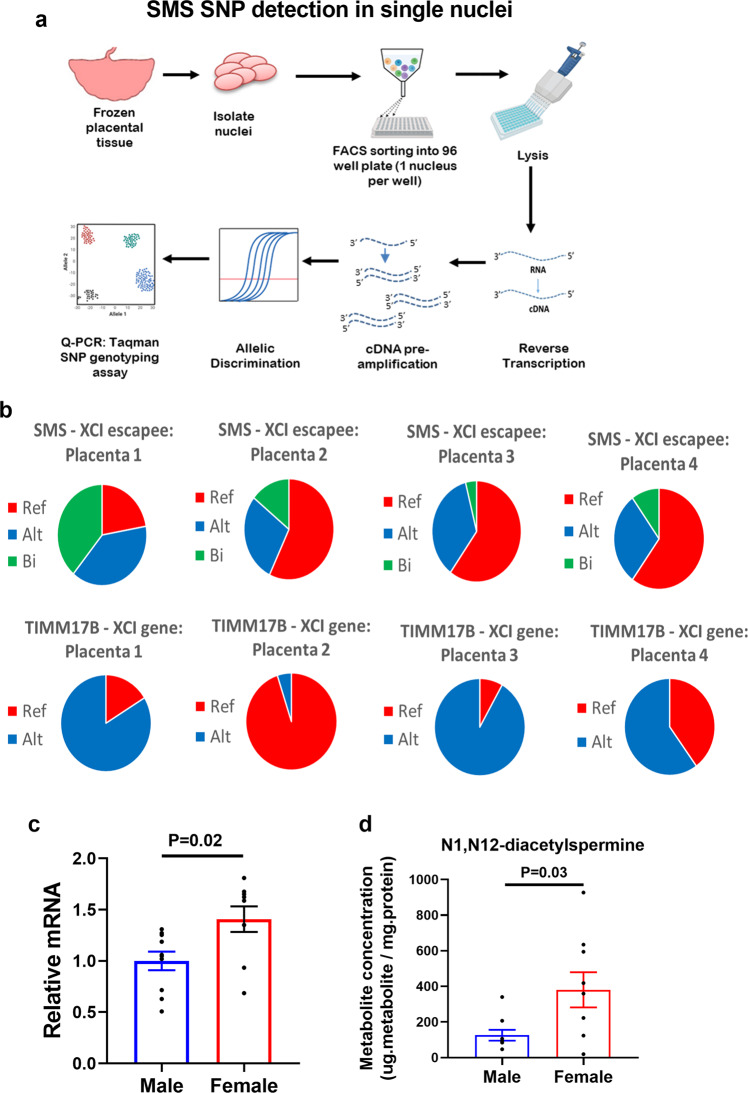
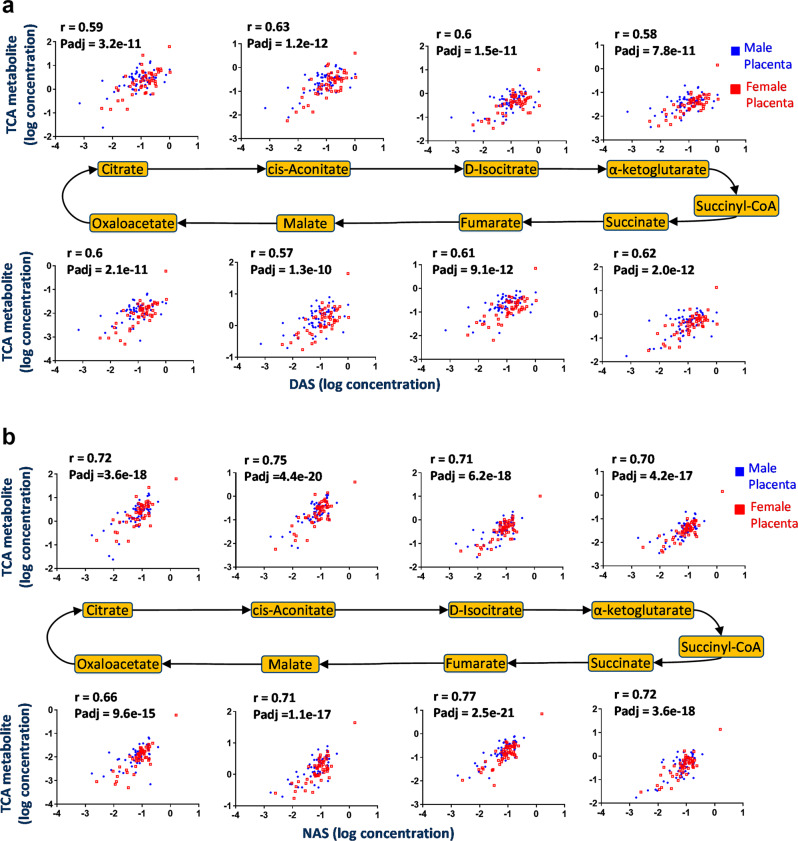
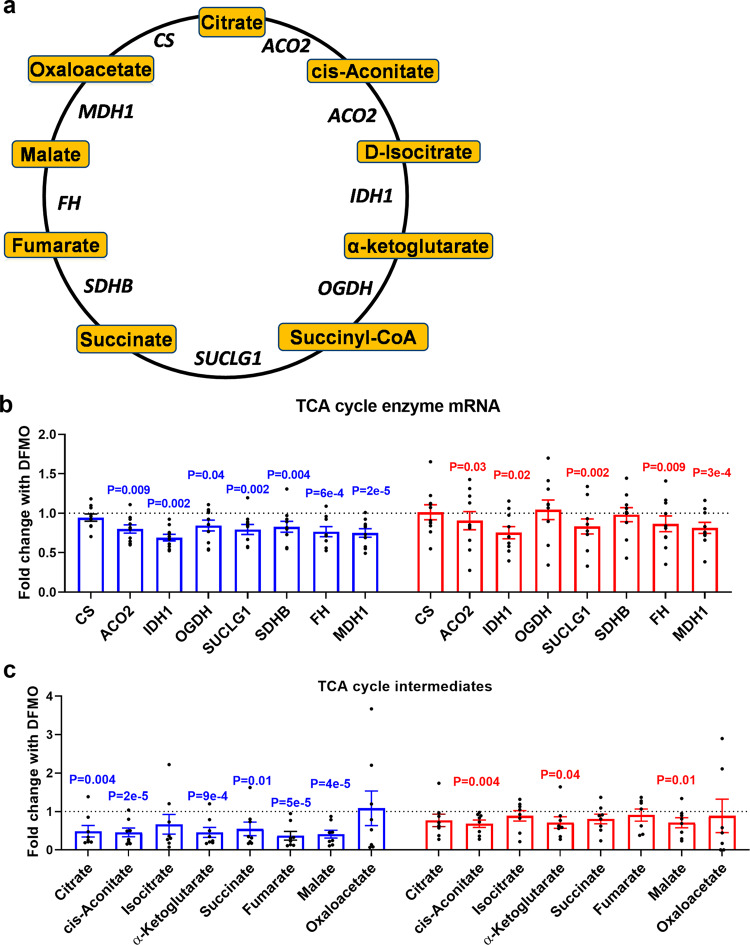
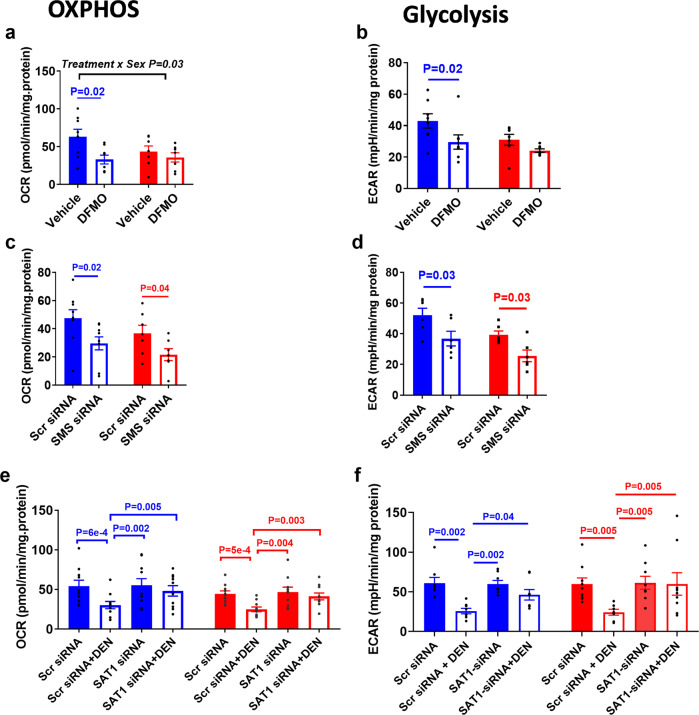
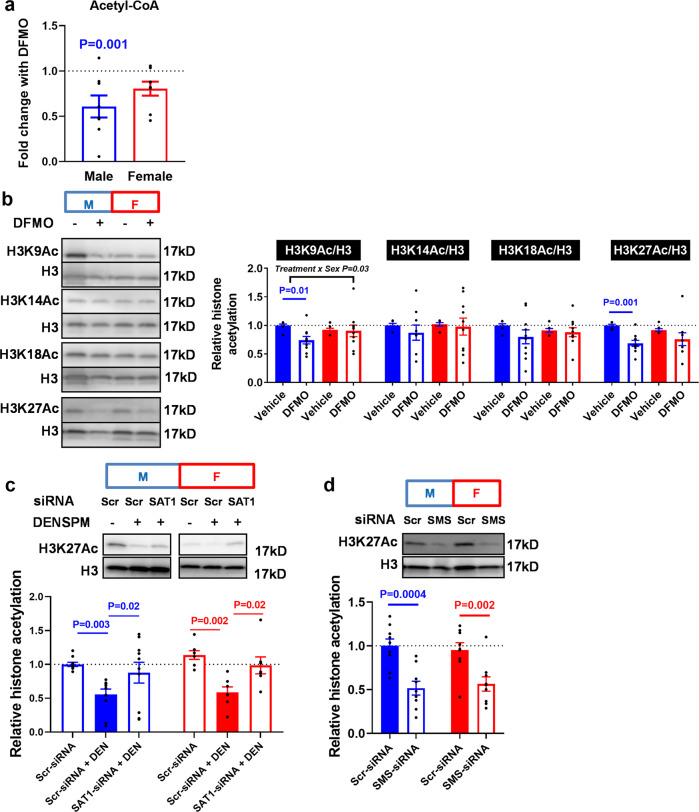
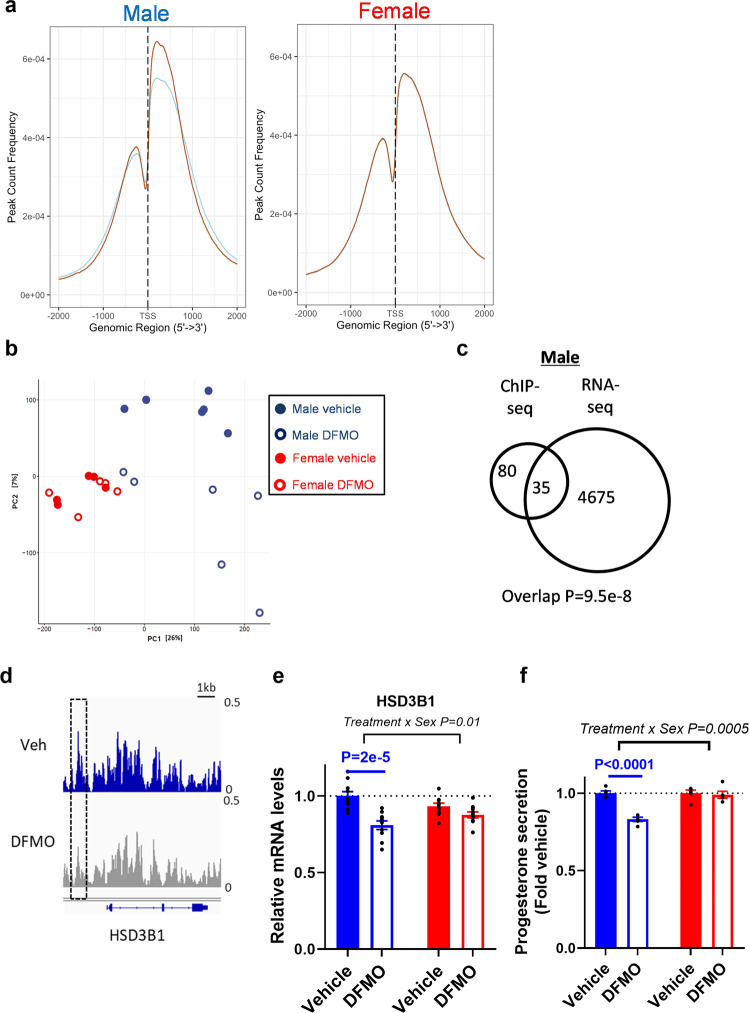
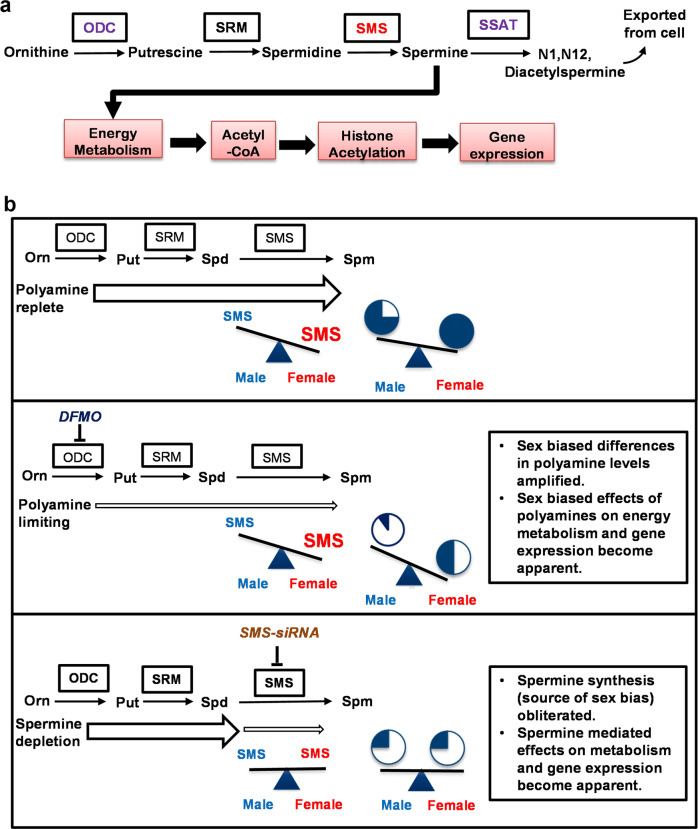
Placental function and dysfunction differ by sex but the mechanisms are unknown. Here we show that sex differences in polyamine metabolism are associated with escape from X chromosome inactivation of the gene encoding spermine synthase (SMS). Female placental trophoblasts demonstrate biallelic SMS expression, associated with increased SMS mRNA and enzyme activity. Polyamine depletion in primary trophoblasts reduced glycolysis and oxidative phosphorylation resulting in decreased acetyl-coA availability and global histone hypoacetylation in a sex-dependent manner. Chromatin-immunoprecipitation sequencing and RNA-sequencing identifies progesterone biosynthesis as a target of polyamine regulated gene expression, and polyamine depletion reduced progesterone release in male trophoblasts. The effects of polyamine depletion can be attributed to spermine as SMS-silencing recapitulated the effects on energy metabolism, histone acetylation, and progesterone release. In summary, spermine metabolism alters trophoblast gene expression through acetyl-coA biosynthesis and histone acetylation, and SMS escape from X inactivation explains some features of human placental sex differences.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- W.H.O. World Health Organisation: The Global Burden of Disease 2004 Update (W.H.O., 2008).
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
