

S. cerevisiae Cells Can Grow without the Pds5 Cohesin Subunit
- PMID: 35708277
- PMCID: PMC9426526
- DOI: 10.1128/mbio.01420-22
S. cerevisiae Cells Can Grow without the Pds5 Cohesin Subunit
Abstract
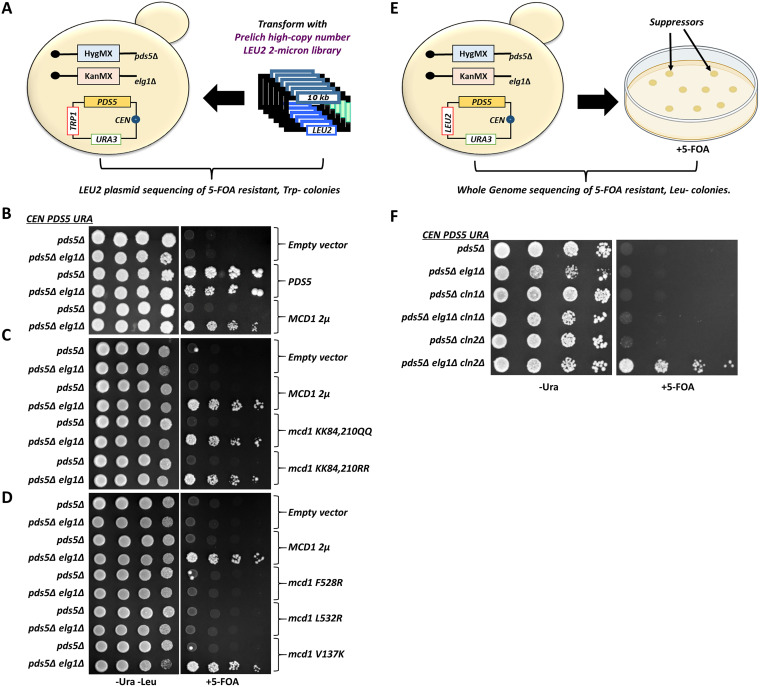
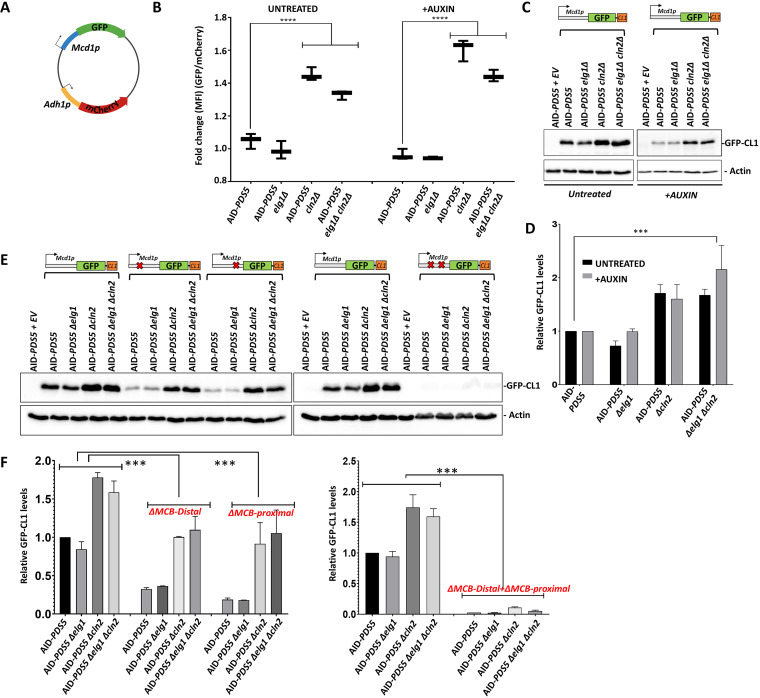
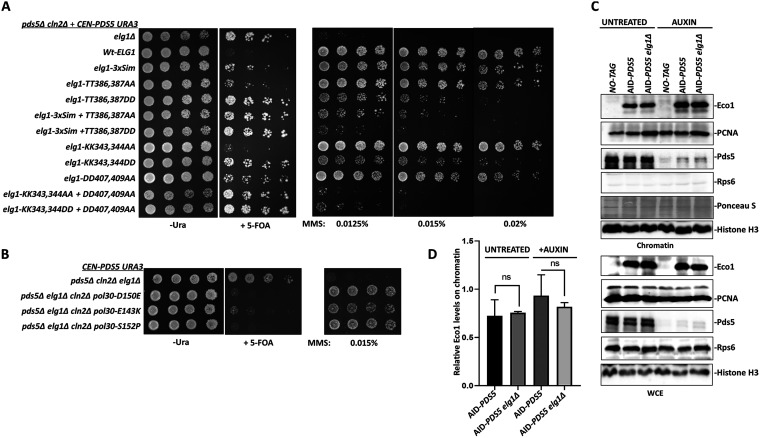
During DNA replication, the newly created sister chromatids are held together until their separation at anaphase. The cohesin complex is in charge of creating and maintaining sister chromatid cohesion (SCC) in all eukaryotes. In Saccharomyces cerevisiae cells, cohesin is composed of two elongated proteins, Smc1 and Smc3, bridged by the kleisin Mcd1/Scc1. The latter also acts as a scaffold for three additional proteins, Scc3/Irr1, Wpl1/Rad61, and Pds5. Although the HEAT-repeat protein Pds5 is essential for cohesion, its precise function is still debated. Deletion of the ELG1 gene, encoding a PCNA unloader, can partially suppress the temperature-sensitive pds5-1 allele, but not a complete deletion of PDS5. We carried out a genetic screen for high-copy-number suppressors and another for spontaneously arising mutants, allowing the survival of a pds5Δ elg1Δ strain. Our results show that cells remain viable in the absence of Pds5 provided that there is both an elevation in the level of Mcd1 (which can be due to mutations in the CLN2 gene, encoding a G1 cyclin), and an increase in the level of SUMO-modified PCNA on chromatin (caused by lack of PCNA unloading in elg1Δ mutants). The elevated SUMO-PCNA levels increase the recruitment of the Srs2 helicase, which evicts Rad51 molecules from the moving fork, creating single-stranded DNA (ssDNA) regions that serve as sites for increased cohesin loading and SCC establishment. Thus, our results delineate a double role for Pds5 in protecting the cohesin ring and interacting with the DNA replication machinery. IMPORTANCE Sister chromatid cohesion is vital for faithful chromosome segregation, chromosome folding into loops, and gene expression. A multisubunit protein complex known as cohesin holds the sister chromatids from S phase until the anaphase stage. In this study, we explore the function of the essential cohesin subunit Pds5 in the regulation of sister chromatid cohesion. We performed two independent genetic screens to bypass the function of the Pds5 protein. We observe that Pds5 protein is a cohesin stabilizer, and elevating the levels of Mcd1 protein along with SUMO-PCNA accumulation on chromatin can compensate for the loss of the PDS5 gene. In addition, Pds5 plays a role in coordinating the DNA replication and sister chromatid cohesion establishment. This work elucidates the function of cohesin subunit Pds5, the G1 cyclin Cln2, and replication factors PCNA, Elg1, and Srs2 in the proper regulation of sister chromatid cohesion.
Keywords: Cln2; DNA replication; ELG1; Elg1; Mcd1; PCNA and PDS5; Pds5; Rad51; SUMO; Srs2; cohesin; sister chromatid cohesion; yeast.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




Similar articles
-
Cohesin without cohesion: a novel role for Pds5 in Saccharomyces cerevisiae.PLoS One. 2014 Jun 25;9(6):e100470. doi: 10.1371/journal.pone.0100470. eCollection 2014. PLoS One. 2014. PMID: 24963665 Free PMC article.
-
Cohesin rings devoid of Scc3 and Pds5 maintain their stable association with the DNA.PLoS Genet. 2012;8(8):e1002856. doi: 10.1371/journal.pgen.1002856. Epub 2012 Aug 9. PLoS Genet. 2012. PMID: 22912589 Free PMC article.
-
Pds5 regulators segregate cohesion and condensation pathways in Saccharomyces cerevisiae.Proc Natl Acad Sci U S A. 2015 Jun 2;112(22):7021-6. doi: 10.1073/pnas.1501369112. Epub 2015 May 18. Proc Natl Acad Sci U S A. 2015. PMID: 25986377 Free PMC article.
-
The Interplay of Cohesin and the Replisome at Processive and Stressed DNA Replication Forks.Cells. 2021 Dec 8;10(12):3455. doi: 10.3390/cells10123455. Cells. 2021. PMID: 34943967 Free PMC article. Review.
-
[Sister chromatid cohesion complex in Eukaryota].Postepy Biochem. 2010;56(1):41-54. Postepy Biochem. 2010. PMID: 20499680 Review. Polish.
Cited by
-
It's all in the numbers: Cohesin stoichiometry.Front Mol Biosci. 2022 Oct 18;9:1010894. doi: 10.3389/fmolb.2022.1010894. eCollection 2022. Front Mol Biosci. 2022. PMID: 36330215 Free PMC article. Review.
-
PCNA recruits cohesin loader Scc2 to ensure sister chromatid cohesion.Nat Struct Mol Biol. 2023 Sep;30(9):1286-1294. doi: 10.1038/s41594-023-01064-x. Epub 2023 Aug 17. Nat Struct Mol Biol. 2023. PMID: 37592094 Free PMC article.
-
Fdo1, Fkh1, Fkh2, and the Swi6-Mbp1 MBF complex regulate Mcd1 levels to impact eco1 rad61 cell growth in Saccharomyces cerevisiae.Genetics. 2024 Oct 7;228(2):iyae128. doi: 10.1093/genetics/iyae128. Genetics. 2024. PMID: 39110836 Free PMC article.
-
Effects of Defective Unloading and Recycling of PCNA Revealed by the Analysis of ELG1 Mutants.Int J Mol Sci. 2023 Jan 13;24(2):1568. doi: 10.3390/ijms24021568. Int J Mol Sci. 2023. PMID: 36675081 Free PMC article.
-
The cohesin-associated protein Pds5A governs the meiotic spindle assembly via deubiquitination of Kif5B in oocytes.Sci Adv. 2025 Apr 11;11(15):eadt6159. doi: 10.1126/sciadv.adt6159. Epub 2025 Apr 11. Sci Adv. 2025. PMID: 40215310 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
