

Satellite repeat RNA expression in epithelial ovarian cancer associates with a tumor-immunosuppressive phenotype
- PMID: 35708912
- PMCID: PMC9374379
- DOI: 10.1172/JCI155931
Satellite repeat RNA expression in epithelial ovarian cancer associates with a tumor-immunosuppressive phenotype
Abstract
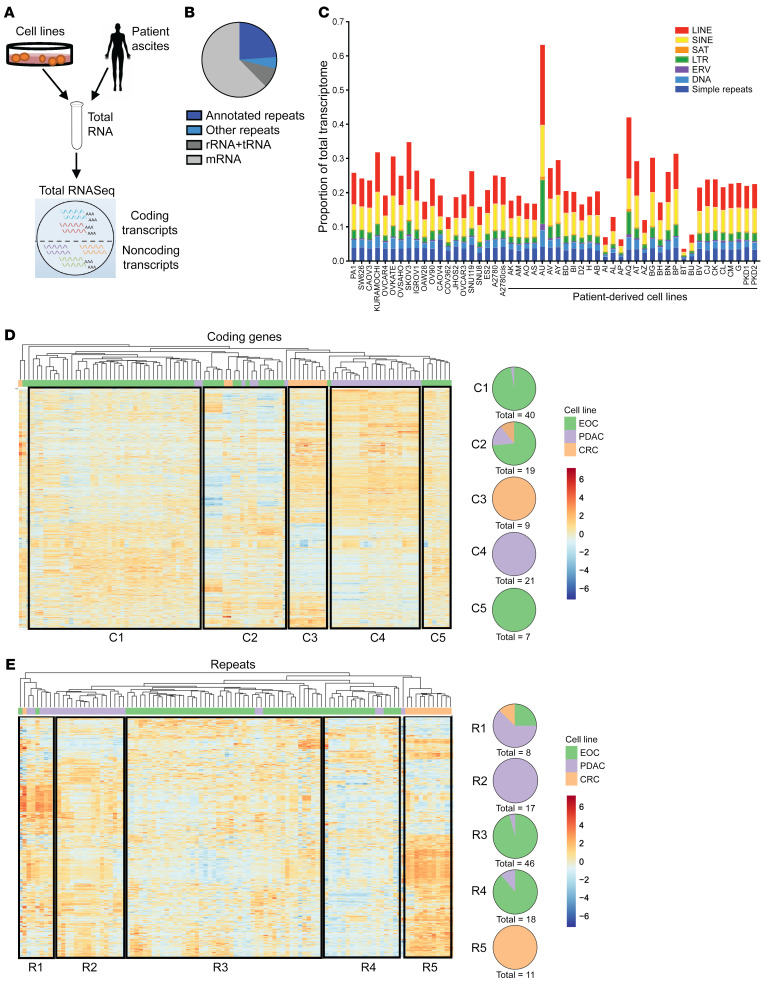
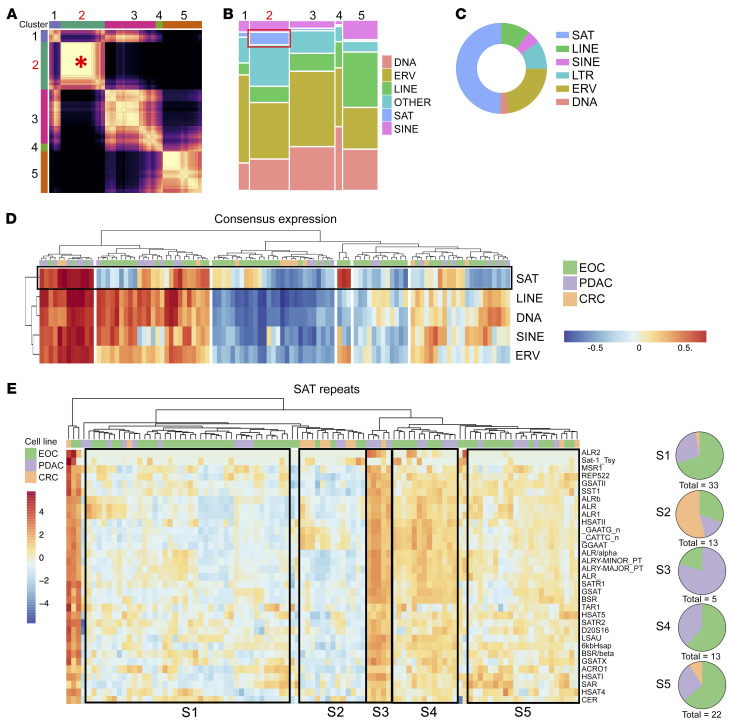
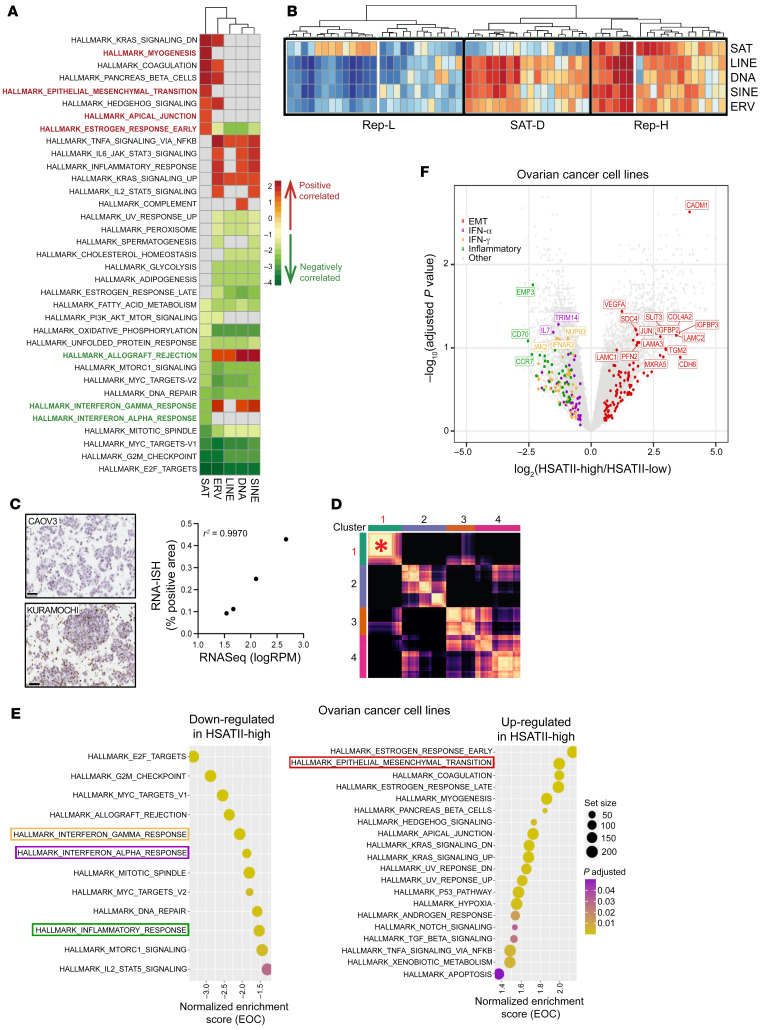
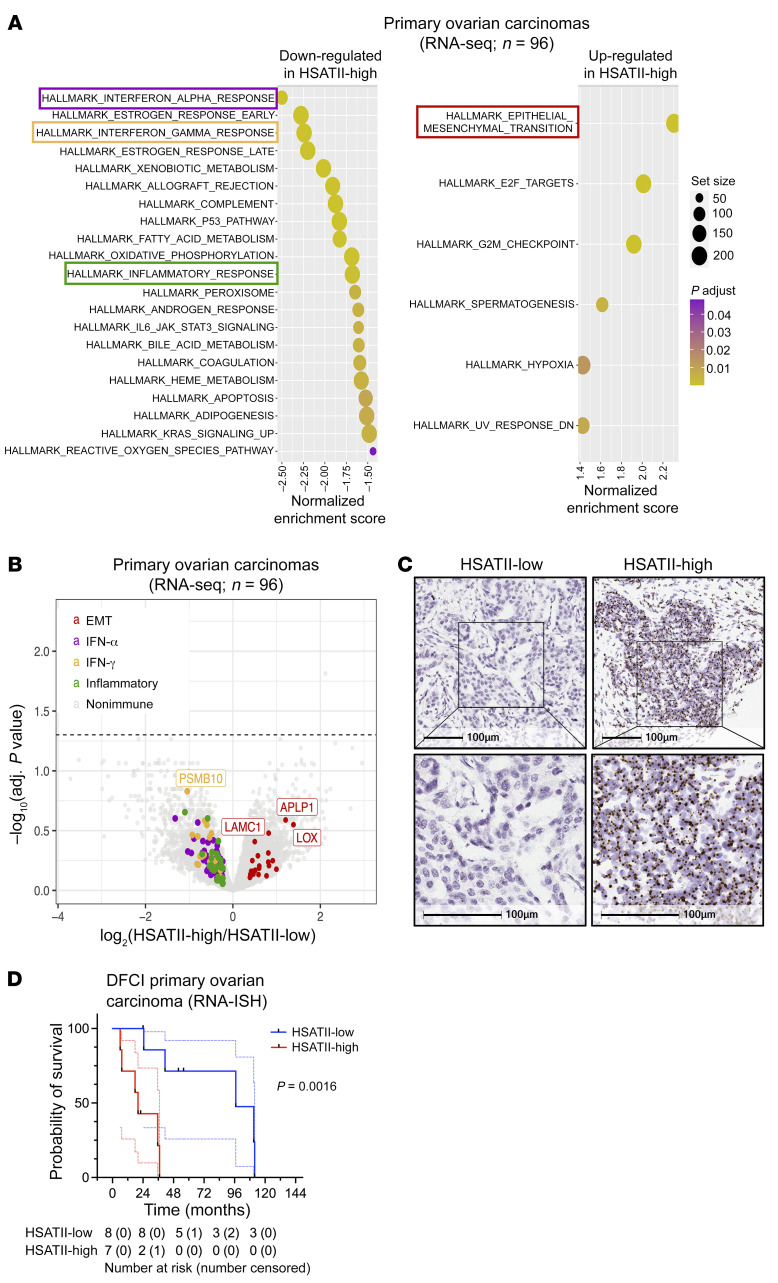
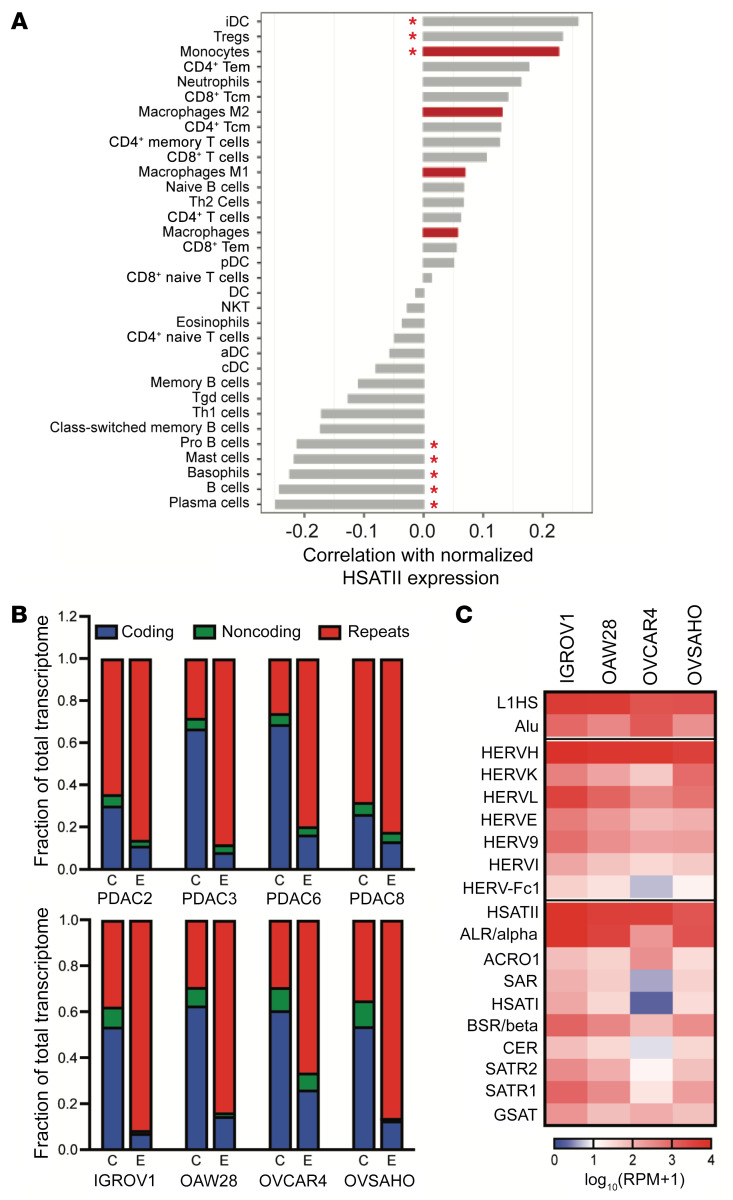
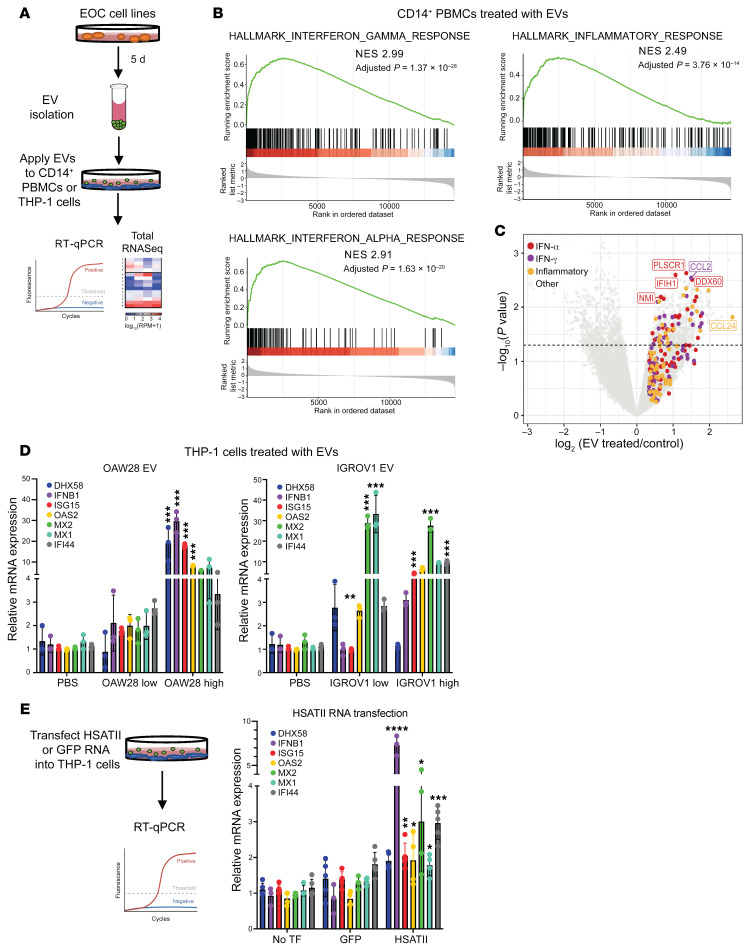
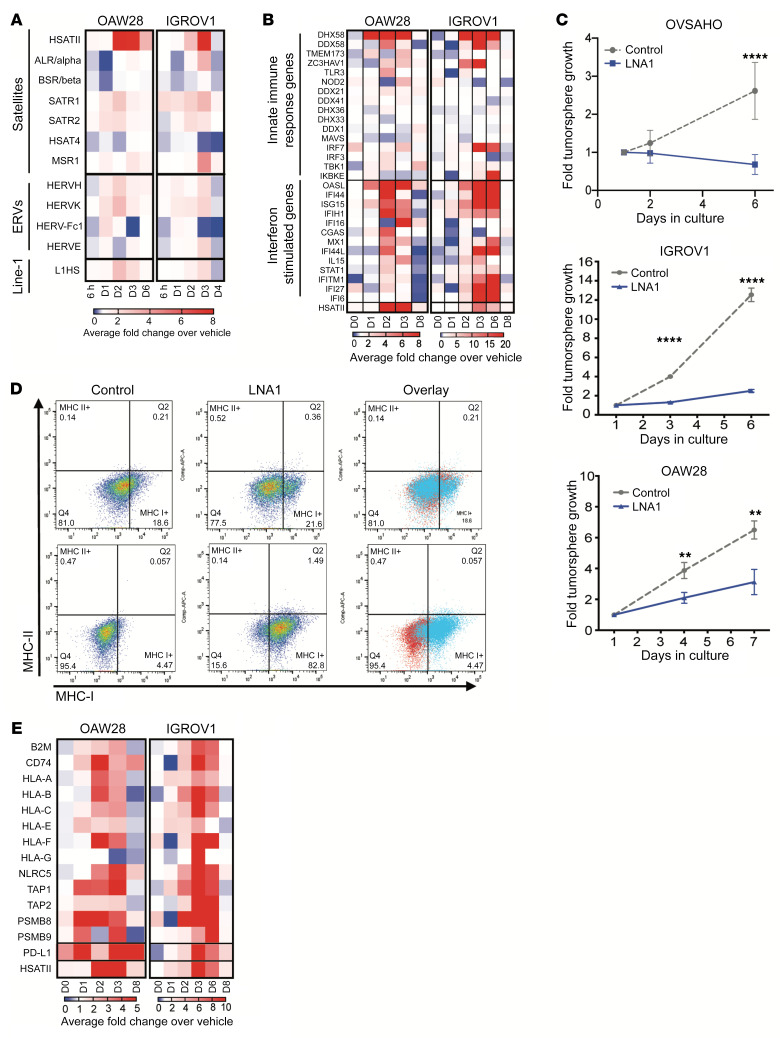
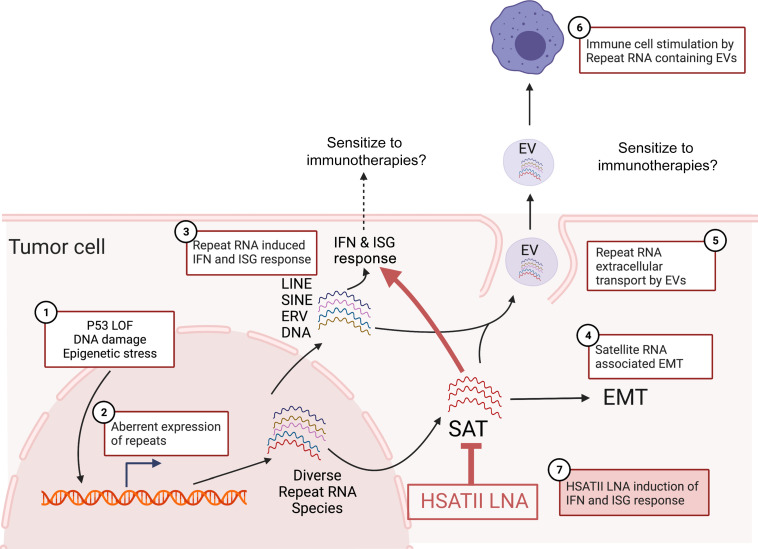
Aberrant expression of viral-like repeat elements is a common feature of epithelial cancers, and the substantial diversity of repeat species provides a distinct view of the cancer transcriptome. Repeatome profiling across ovarian, pancreatic, and colorectal cell lines identifies distinct clustering independent of tissue origin that is seen with coding gene analysis. Deeper analysis of ovarian cancer cell lines demonstrated that human satellite II (HSATII) satellite repeat expression was highly associated with epithelial-mesenchymal transition (EMT) and anticorrelated with IFN-response genes indicative of a more aggressive phenotype. SATII expression - and its correlation with EMT and anticorrelation with IFN-response genes - was also found in ovarian cancer RNA-Seq data and was associated with significantly shorter survival in a second independent cohort of patients with ovarian cancer. Repeat RNAs were enriched in tumor-derived extracellular vesicles capable of stimulating monocyte-derived macrophages, demonstrating a mechanism that alters the tumor microenvironment with these viral-like sequences. Targeting of HSATII with antisense locked nucleic acids stimulated IFN response and induced MHC I expression in ovarian cancer cell lines, highlighting a potential strategy of modulating the repeatome to reestablish antitumor cell immune surveillance.
Keywords: Cancer; Innate immunity; Noncoding RNAs; Oncology.
Figures
Comment in
-
Breaking satellite silence: human satellite II RNA expression in ovarian cancer.J Clin Invest. 2022 Aug 15;132(16):e161981. doi: 10.1172/JCI161981. J Clin Invest. 2022. PMID: 35968784 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
