

Spatial profiling of early primate gastrulation in utero
- PMID: 35709828
- PMCID: PMC7614364
- DOI: 10.1038/s41586-022-04953-1
Spatial profiling of early primate gastrulation in utero
Abstract
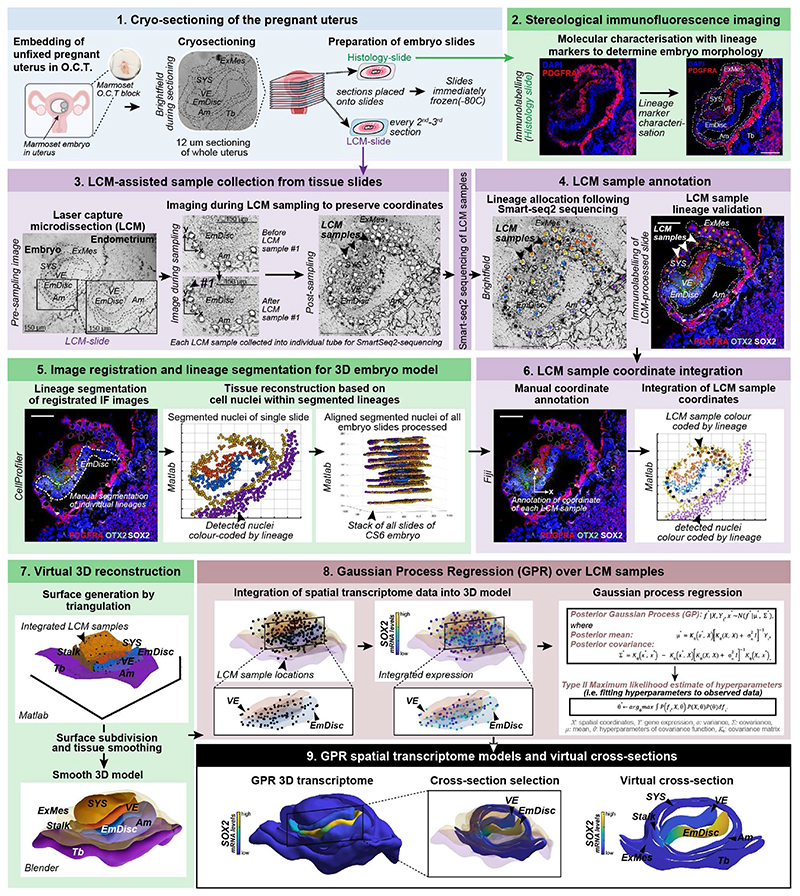
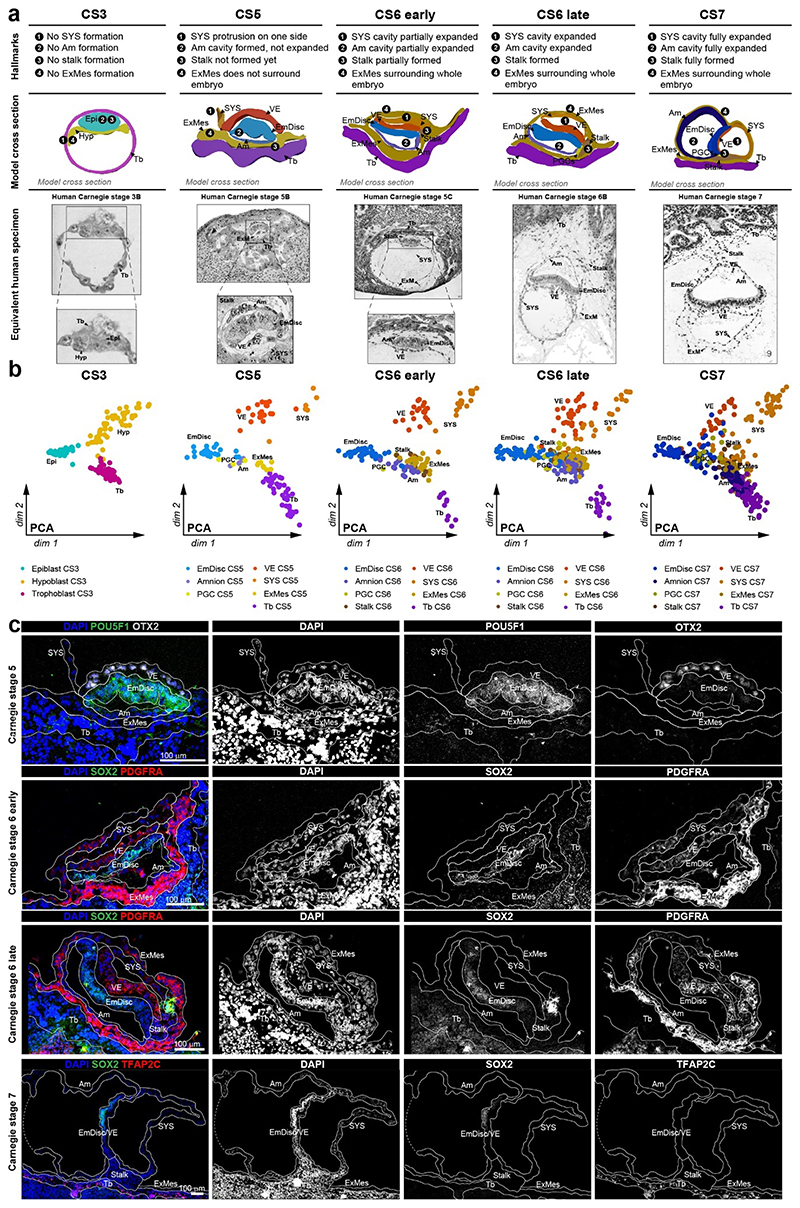
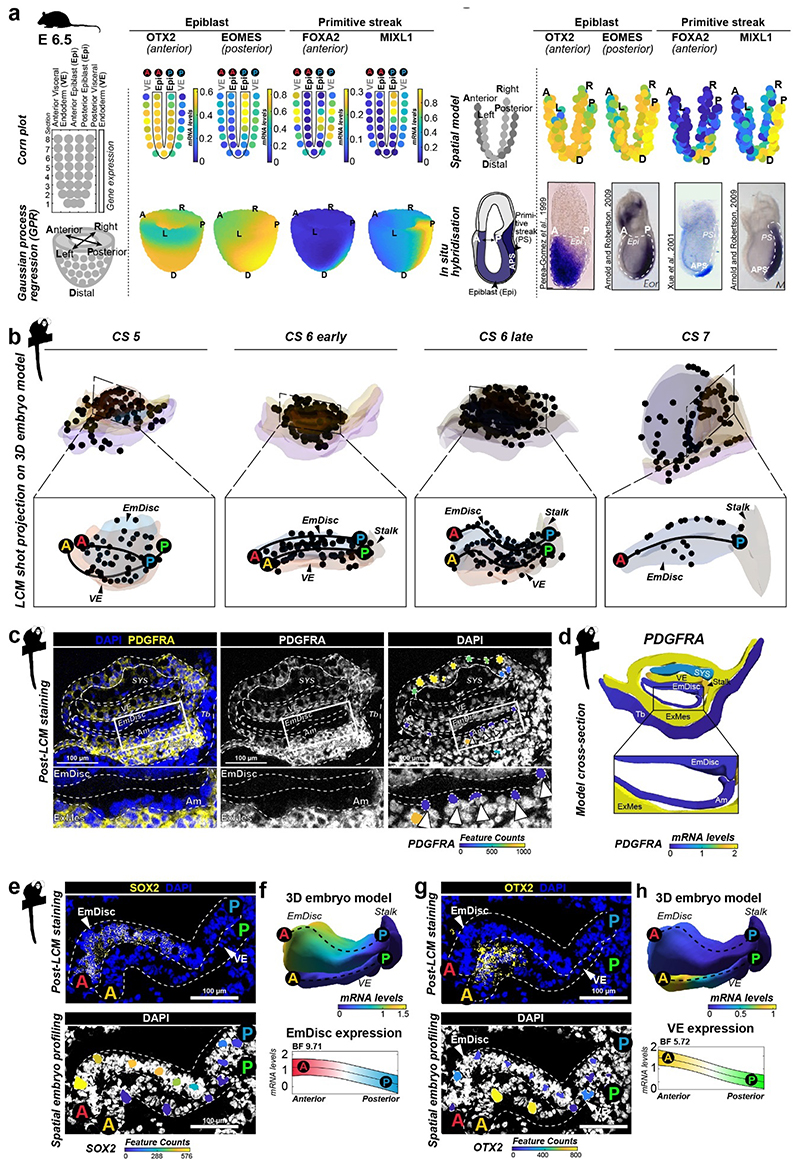
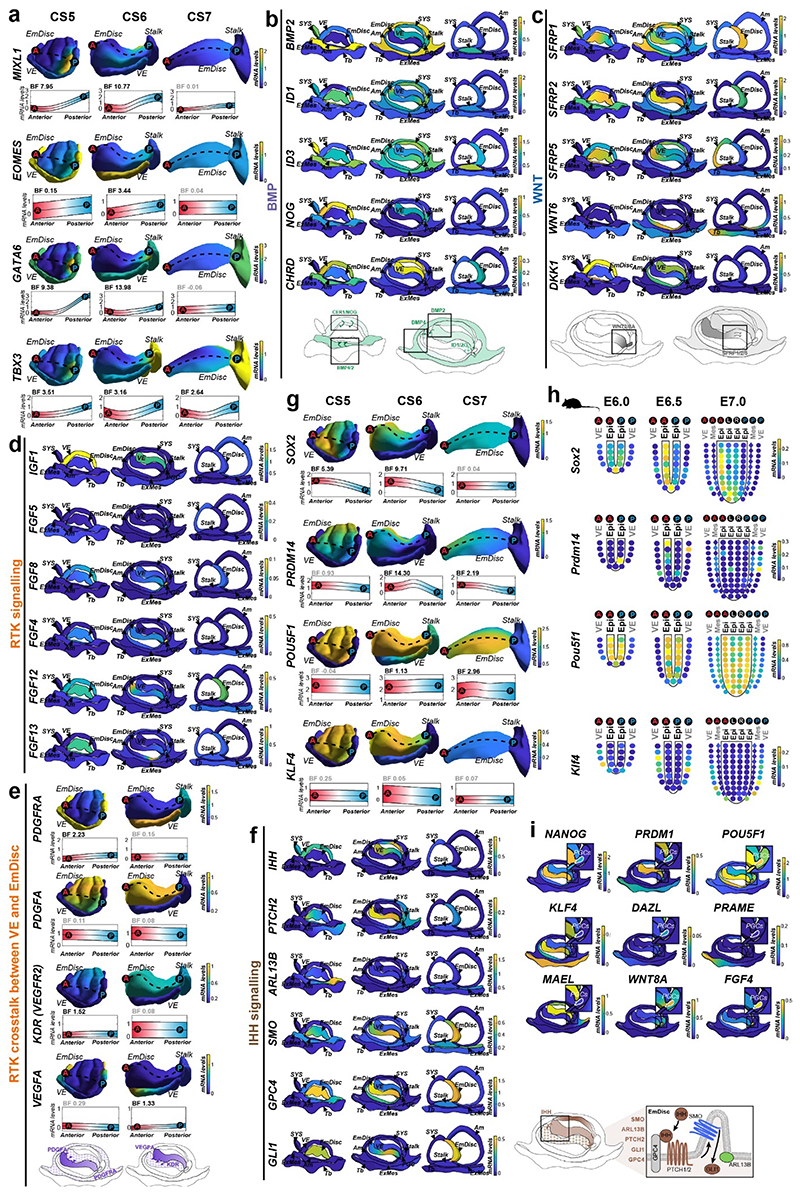
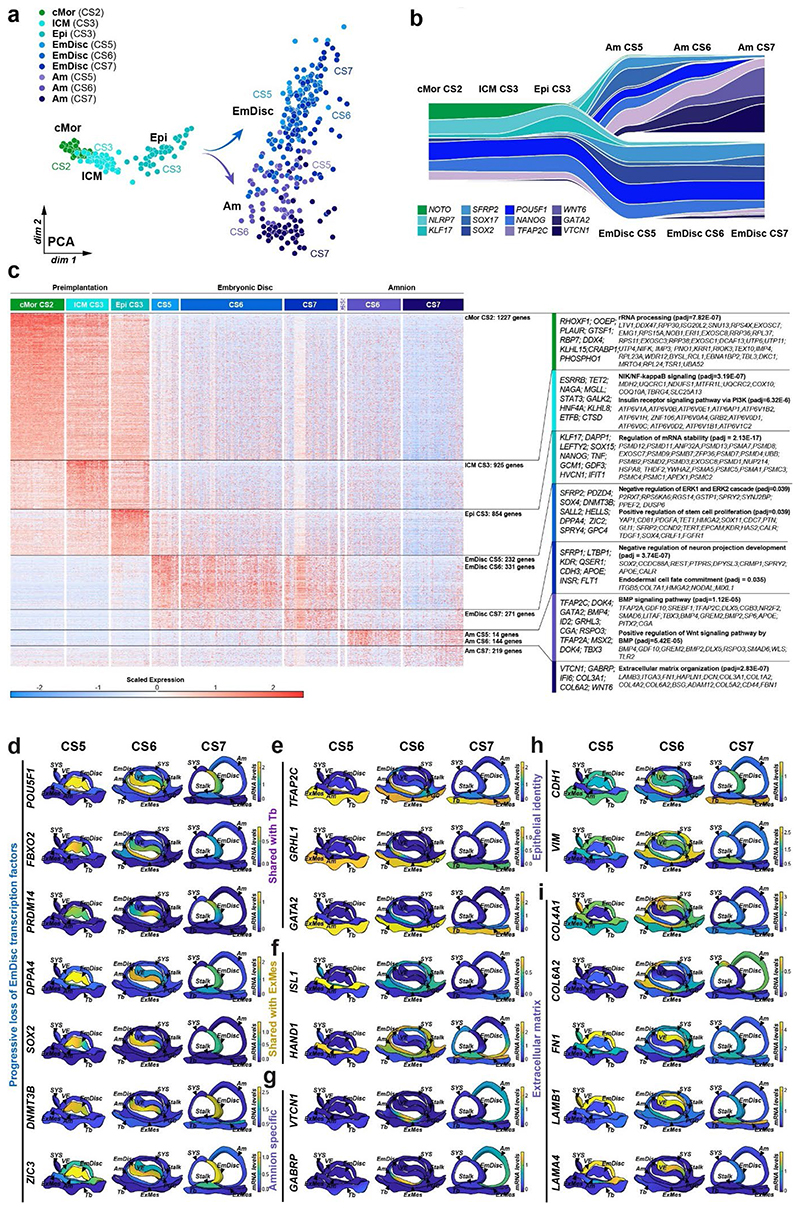
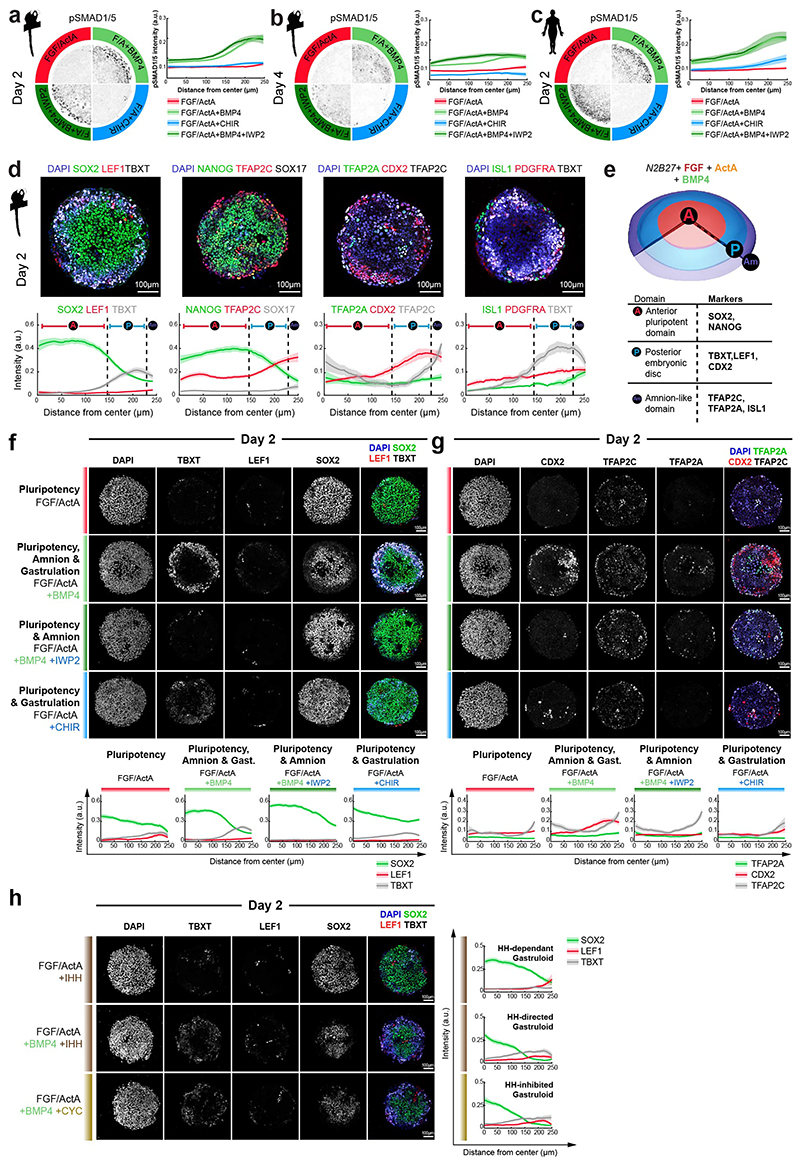
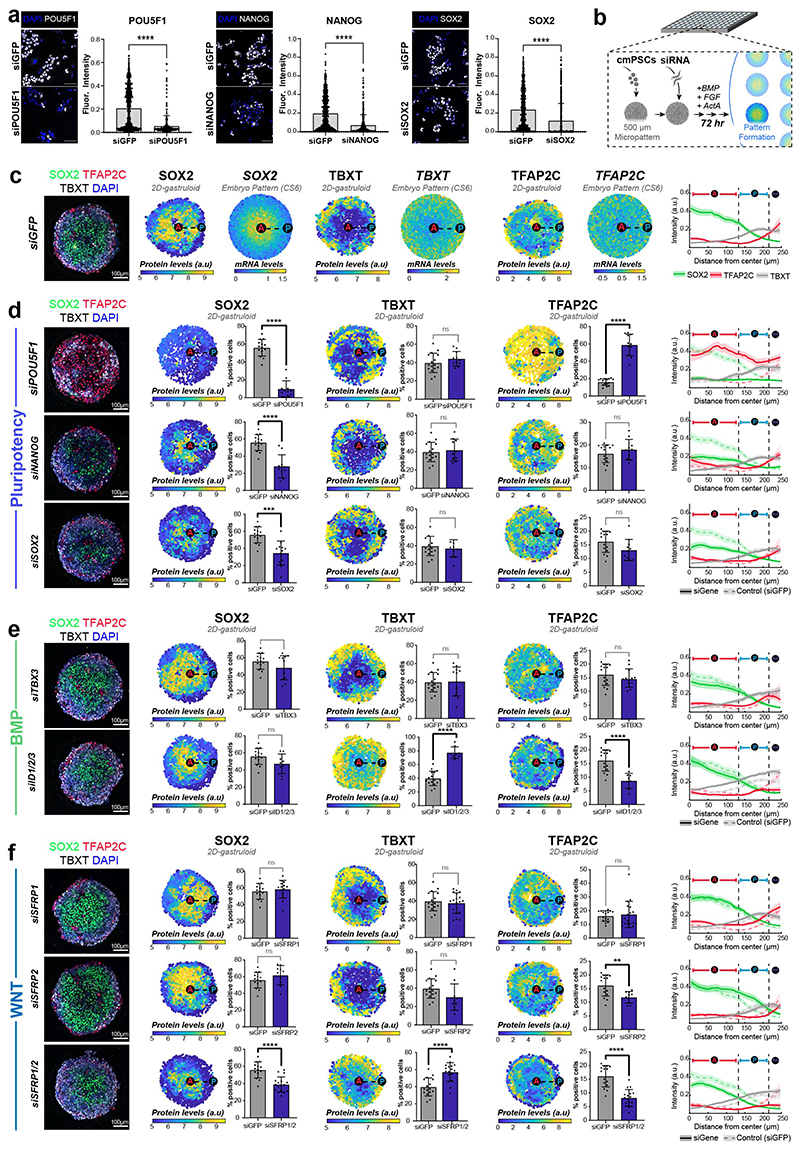
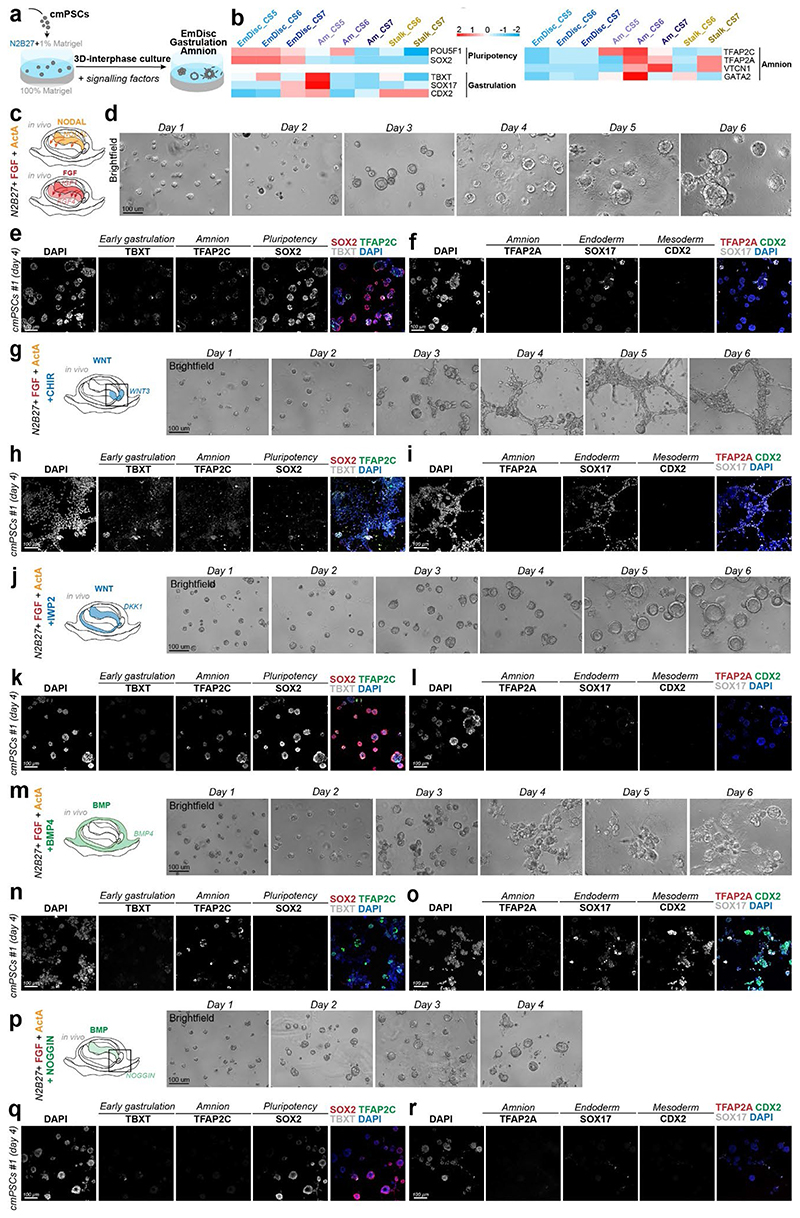
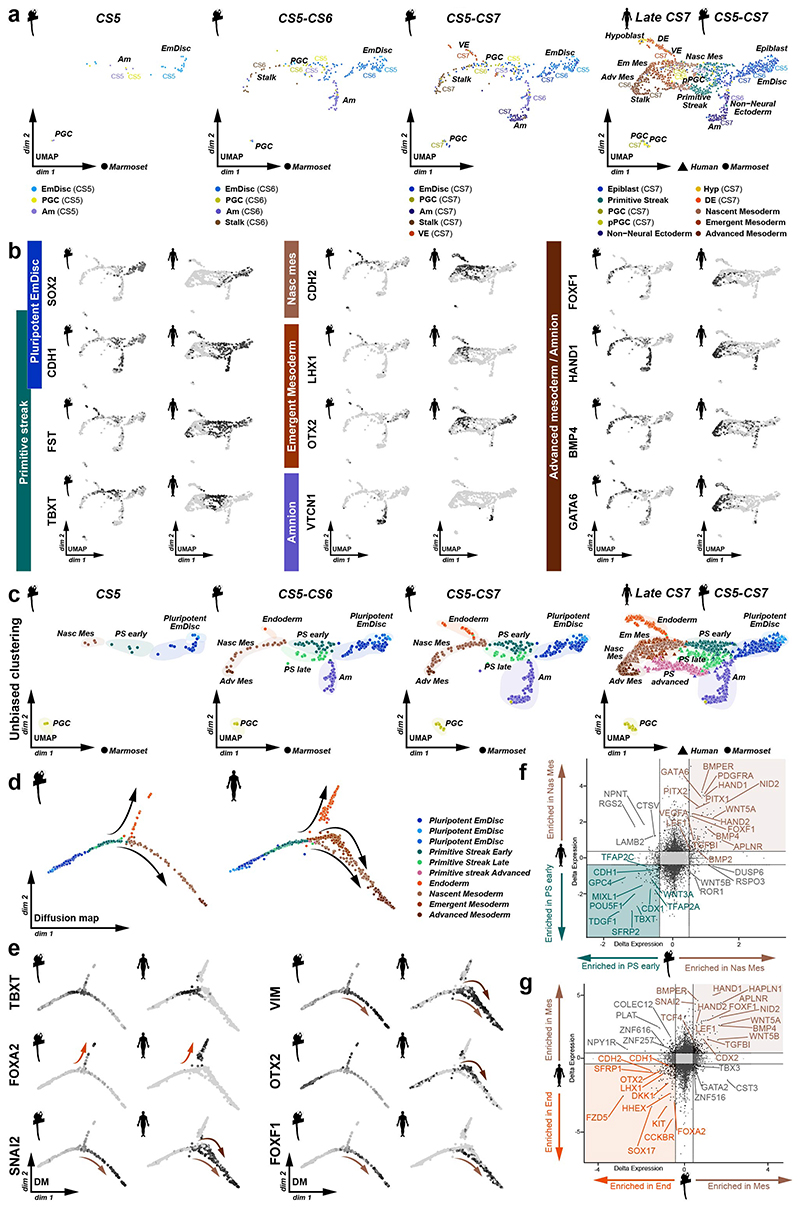
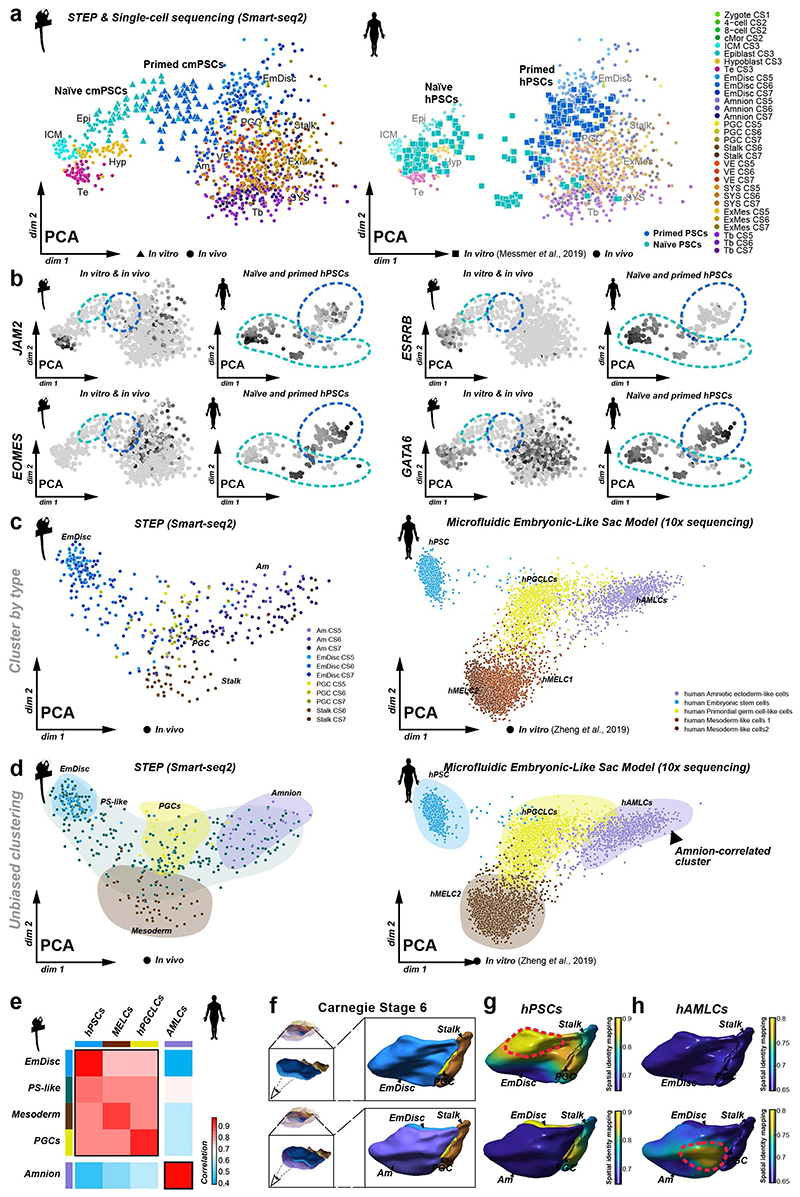
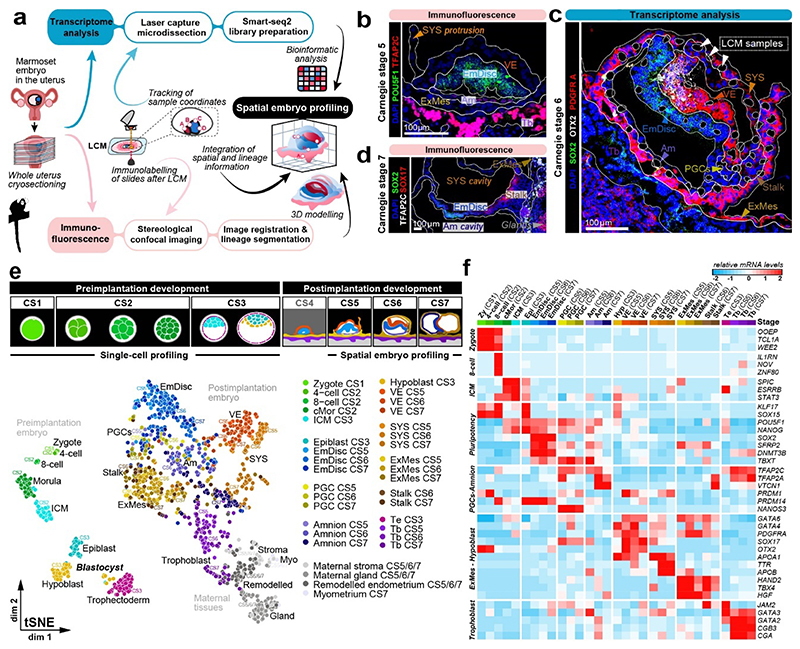
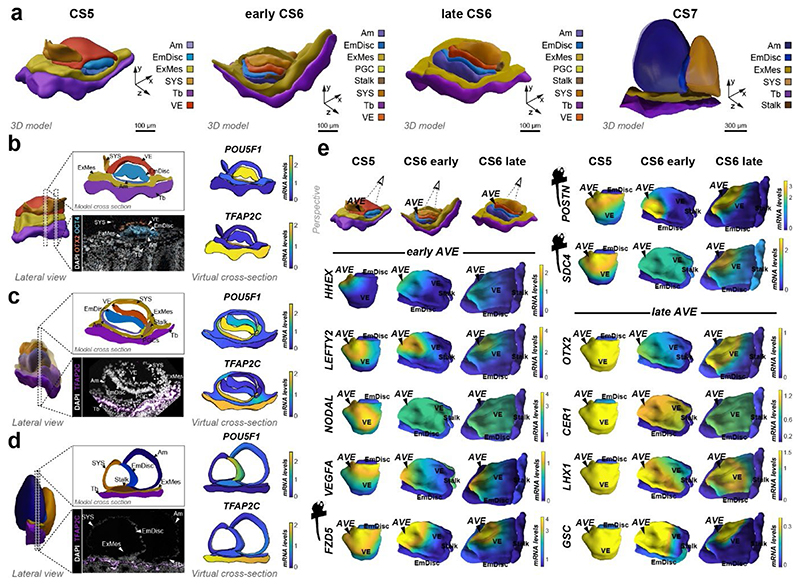
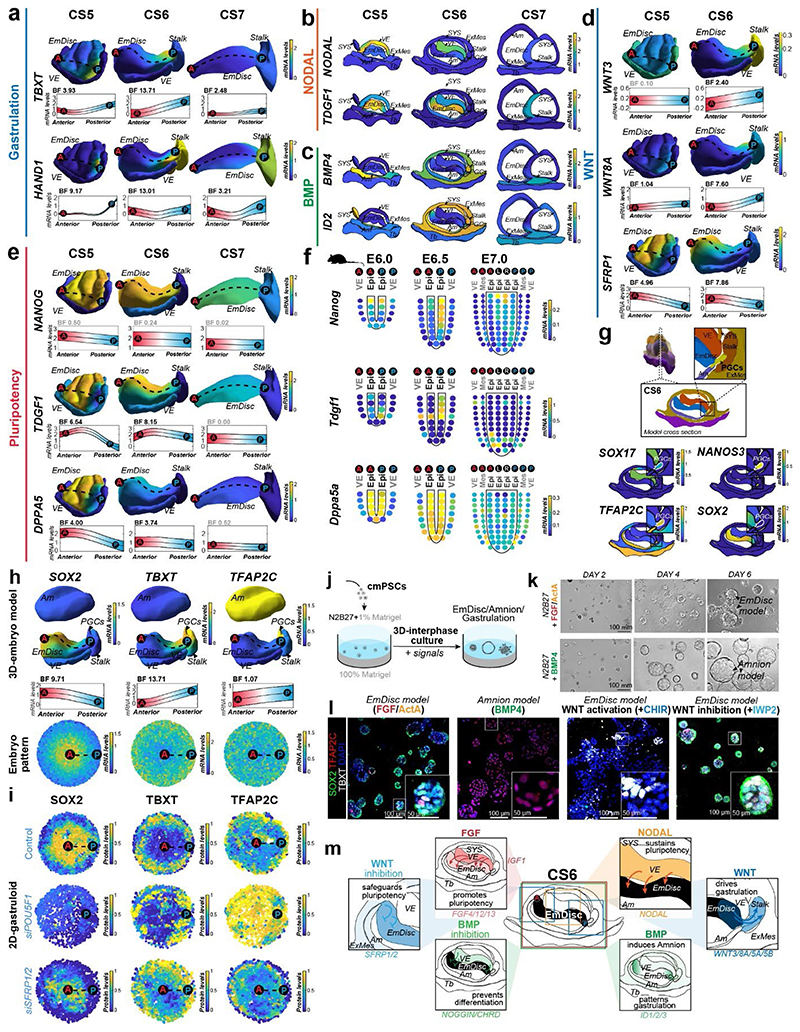
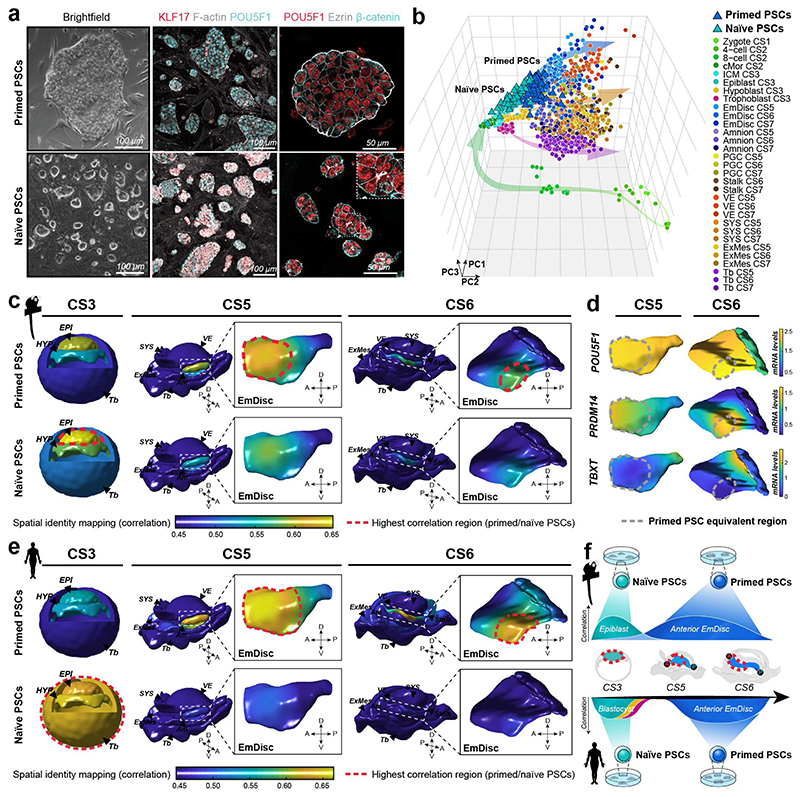
Gastrulation controls the emergence of cellular diversity and axis patterning in the early embryo. In mammals, this transformation is orchestrated by dynamic signalling centres at the interface of embryonic and extraembryonic tissues1-3. Elucidating the molecular framework of axis formation in vivo is fundamental for our understanding of human development4-6 and to advance stem-cell-based regenerative approaches7. Here we illuminate early gastrulation of marmoset embryos in utero using spatial transcriptomics and stem-cell-based embryo models. Gaussian process regression-based 3D transcriptomes delineate the emergence of the anterior visceral endoderm, which is hallmarked by conserved (HHEX, LEFTY2, LHX1) and primate-specific (POSTN, SDC4, FZD5) factors. WNT signalling spatially coordinates the formation of the primitive streak in the embryonic disc and is counteracted by SFRP1 and SFRP2 to sustain pluripotency in the anterior domain. Amnion specification occurs at the boundaries of the embryonic disc through ID1, ID2 and ID3 in response to BMP signalling, providing a developmental rationale for amnion differentiation of primate pluripotent stem cells (PSCs). Spatial identity mapping demonstrates that primed marmoset PSCs exhibit the highest similarity to the anterior embryonic disc, whereas naive PSCs resemble the preimplantation epiblast. Our 3D transcriptome models reveal the molecular code of lineage specification in the primate embryo and provide an in vivo reference to decipher human development.
© 2022. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Tam PP, Loebel DA. Gene function in mouse embryogenesis: get set for gastrulation. Nat Rev Genet. 2007;8:368–381. - PubMed
-
- Rossant J, Tam PPLL. Blastocyst lineage formation, early embryonic asymmetries and axis patterning in the mouse. Development. 2009;136:701–713. - PubMed
-
- Arnold SJ, Robertson EJ. Making a commitment: cell lineage allocation and axis patterning in the early mouse embryo. Nat Rev Mol Cell Biol. 2009;10:91–103. - PubMed
-
- Rossant J, Tam PPL. Exploring early human embryo development. Science (80-.) 2018;360:1075–1076. - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
