

AMH Regulation by Steroids in the Mammalian Testis: Underlying Mechanisms and Clinical Implications
- PMID: 35712256
- PMCID: PMC9195137
- DOI: 10.3389/fendo.2022.906381
AMH Regulation by Steroids in the Mammalian Testis: Underlying Mechanisms and Clinical Implications
Abstract
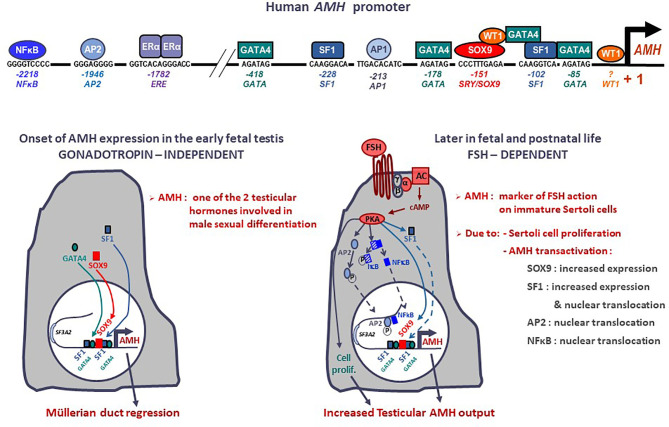
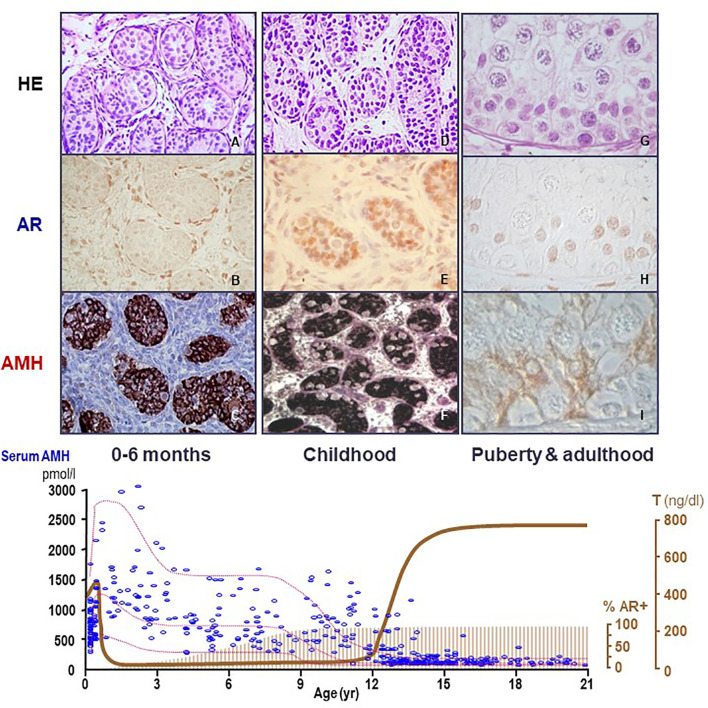
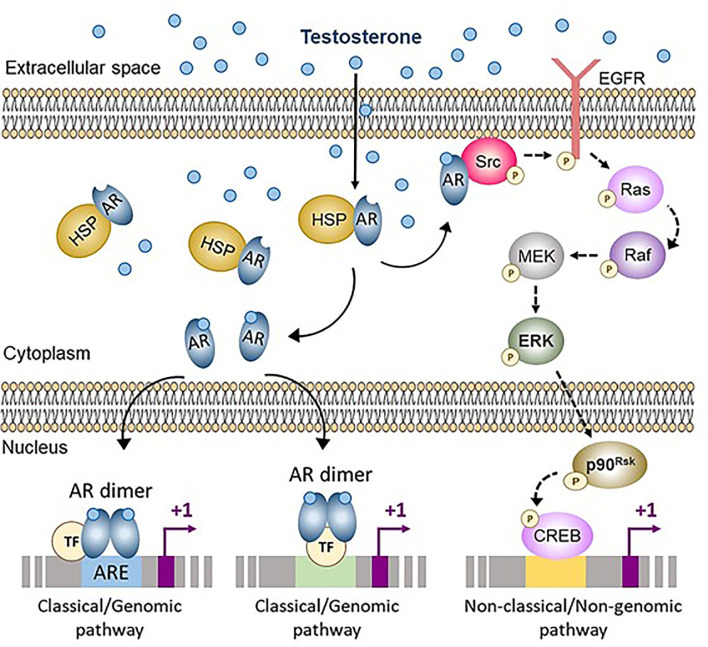
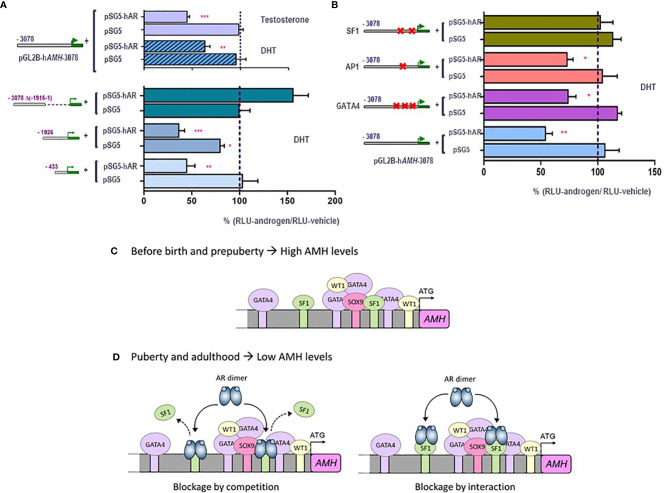
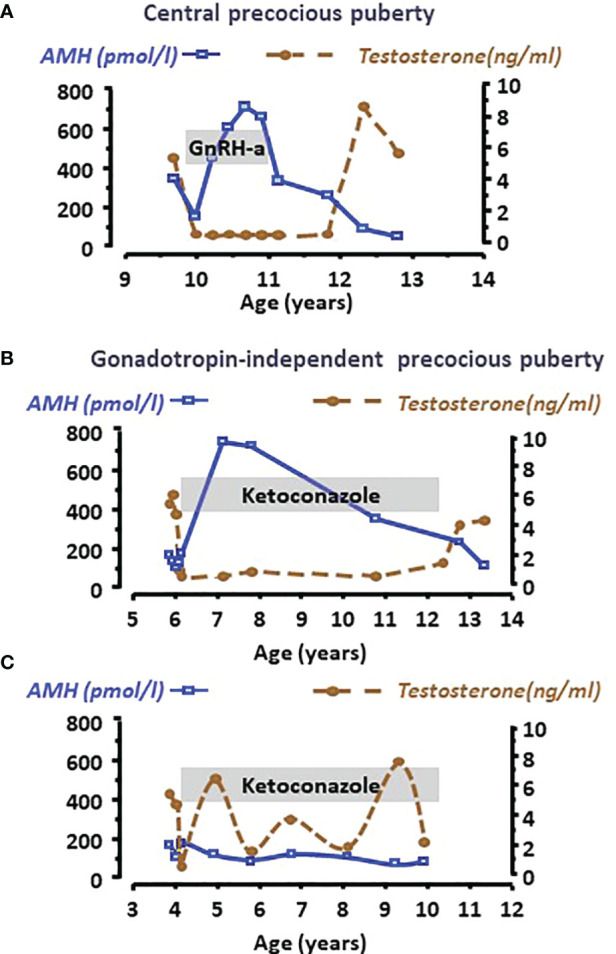
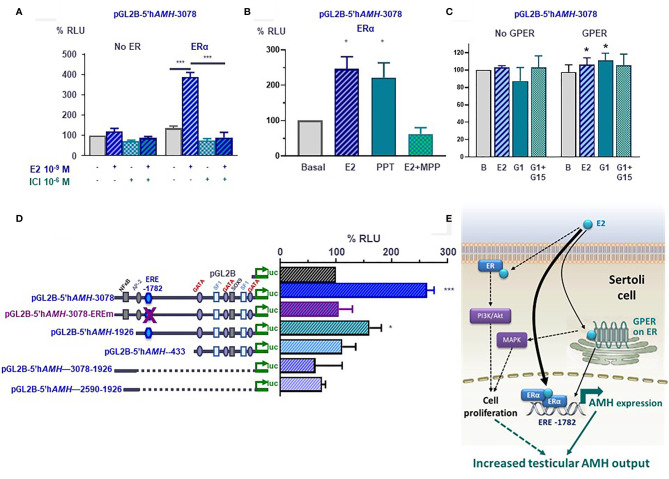
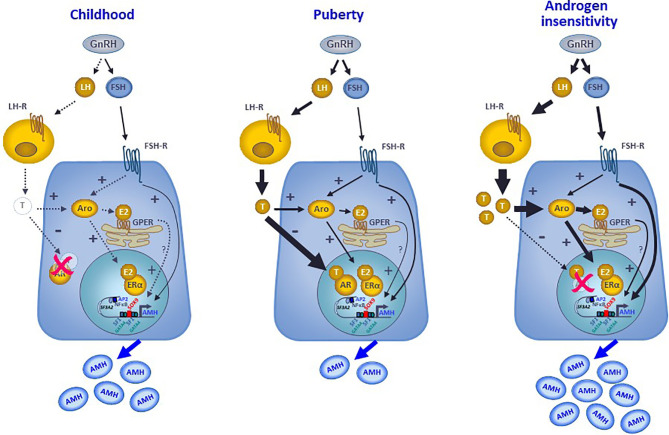
Anti-Müllerian hormone (AMH) is a distinctive biomarker of the immature Sertoli cell. AMH expression, triggered by specific transcription factors upon fetal Sertoli cells differentiation independently of gonadotropins or sex steroids, drives Müllerian duct regression in the male, preventing the development of the uterus and Fallopian tubes. AMH continues to be highly expressed by Sertoli until the onset of puberty, when it is downregulated to low adult levels. FSH increases testicular AMH output by promoting immature Sertoli cell proliferation and individual cell expression. AMH secretion also showcases a differential regulation exerted by intratesticular levels of androgens and estrogens. In the fetus and the newborn, Sertoli cells do not express the androgen receptor, and the high androgen concentrations do not affect AMH expression. Conversely, estrogens can stimulate AMH production because estrogen receptors are present in Sertoli cells and aromatase is stimulated by FSH. During childhood, sex steroids levels are very low and do not play a physiological role on AMH production. However, hyperestrogenic states upregulate AMH expression. During puberty, testosterone inhibition of AMH expression overrides stimulation by estrogens and FSH. The direct effects of sex steroids on AMH transcription are mediated by androgen receptor and estrogen receptor α action on AMH promoter sequences. A modest estrogen action is also mediated by the membrane G-coupled estrogen receptor GPER. The understanding of these complex regulatory mechanisms helps in the interpretation of serum AMH levels found in physiological or pathological conditions, which underscores the importance of serum AMH as a biomarker of intratesticular steroid concentrations.
Keywords: androgen insensitivity; androgen response element; dihydrotestosterone; estradiol; estrogen response element; ketoconazole; testotoxicosis; triptorelin.
Copyright © 2022 Edelsztein, Valeri, Lovaisa, Schteingart and Rey.
Conflict of interest statement
Until 2020, RR received royalties for the development of an AMH ELISA kit and honoraria for technology services using the AMH ELISA. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources