

Complex regulation of Gephyrin splicing is a determinant of inhibitory postsynaptic diversity
- PMID: 35717442
- PMCID: PMC9206673
- DOI: 10.1038/s41467-022-31264-w
Complex regulation of Gephyrin splicing is a determinant of inhibitory postsynaptic diversity
Abstract
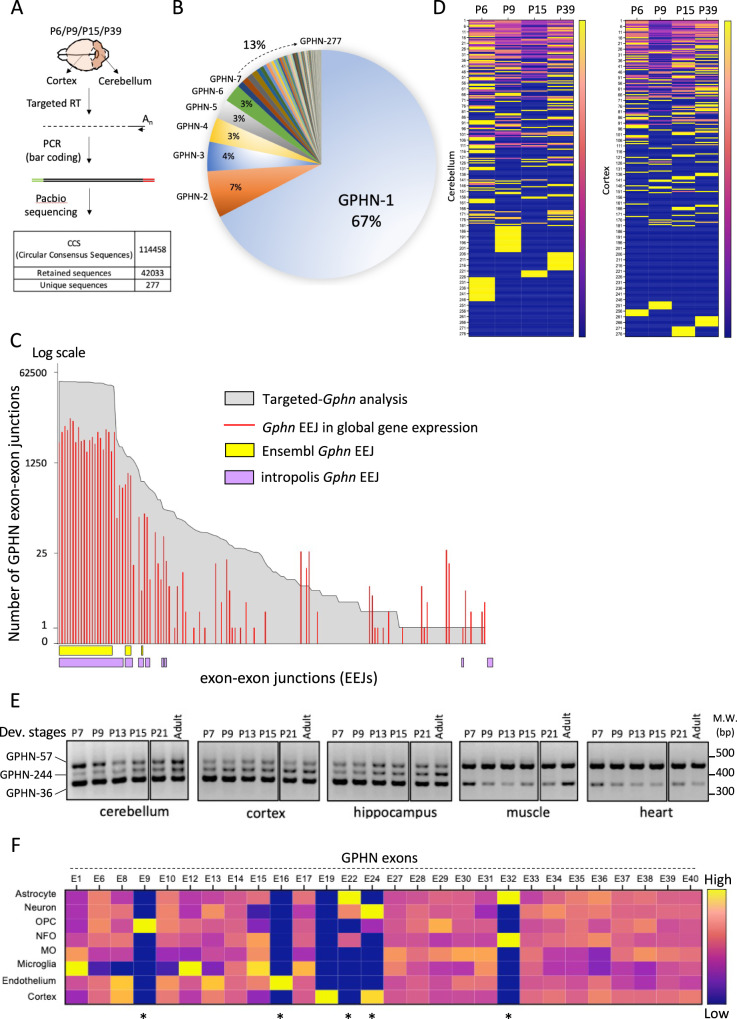
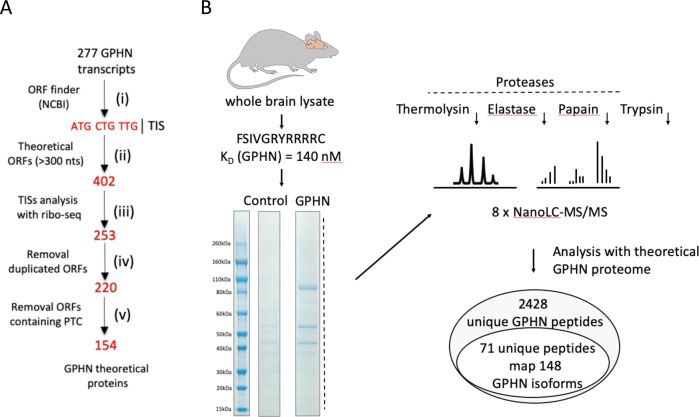
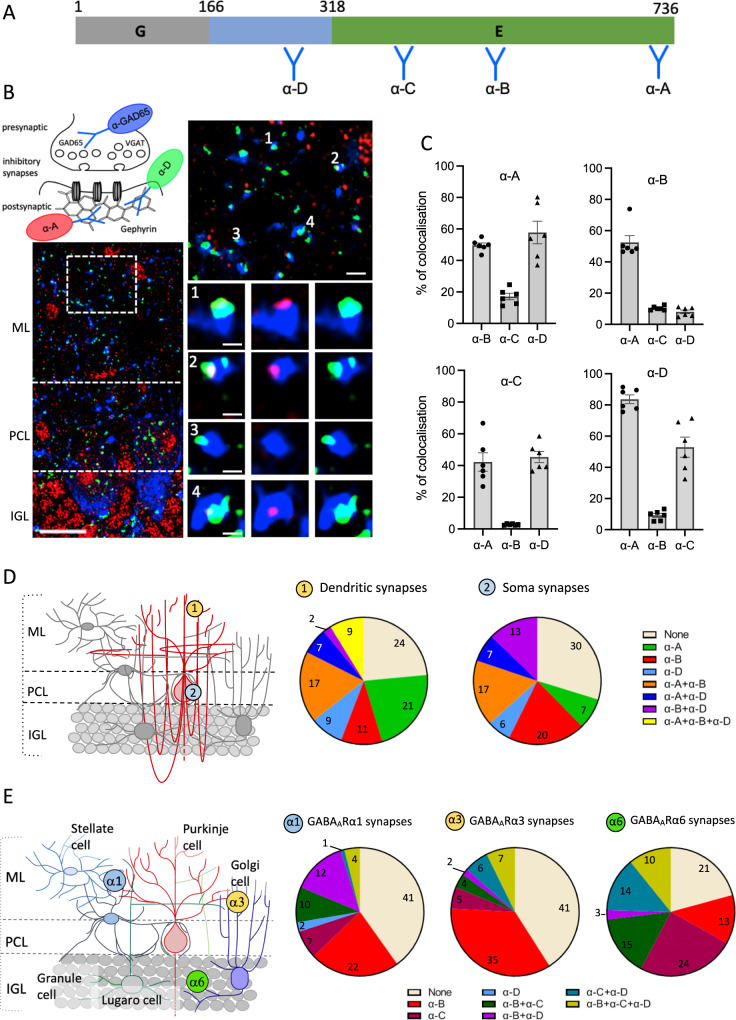
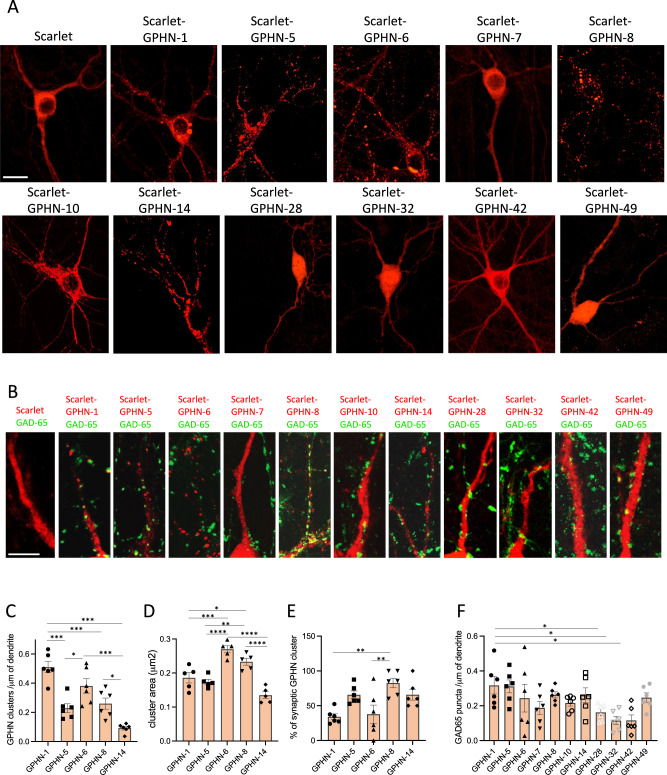
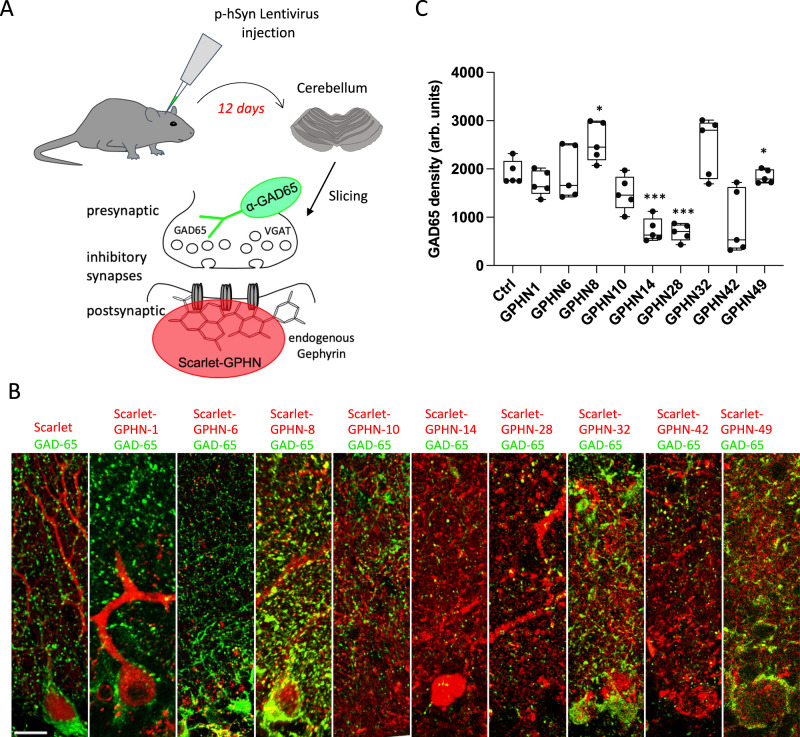
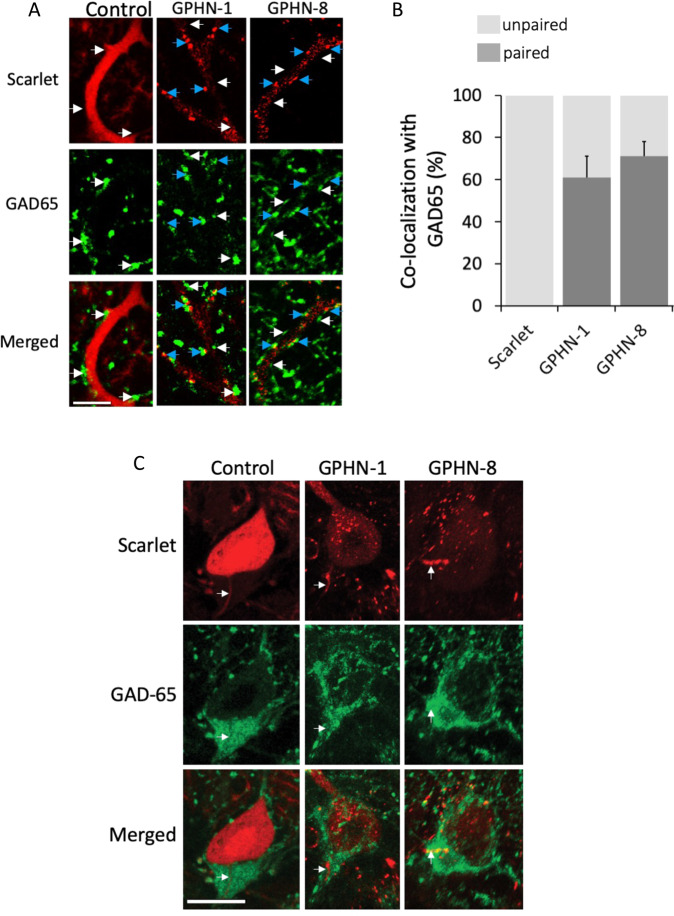
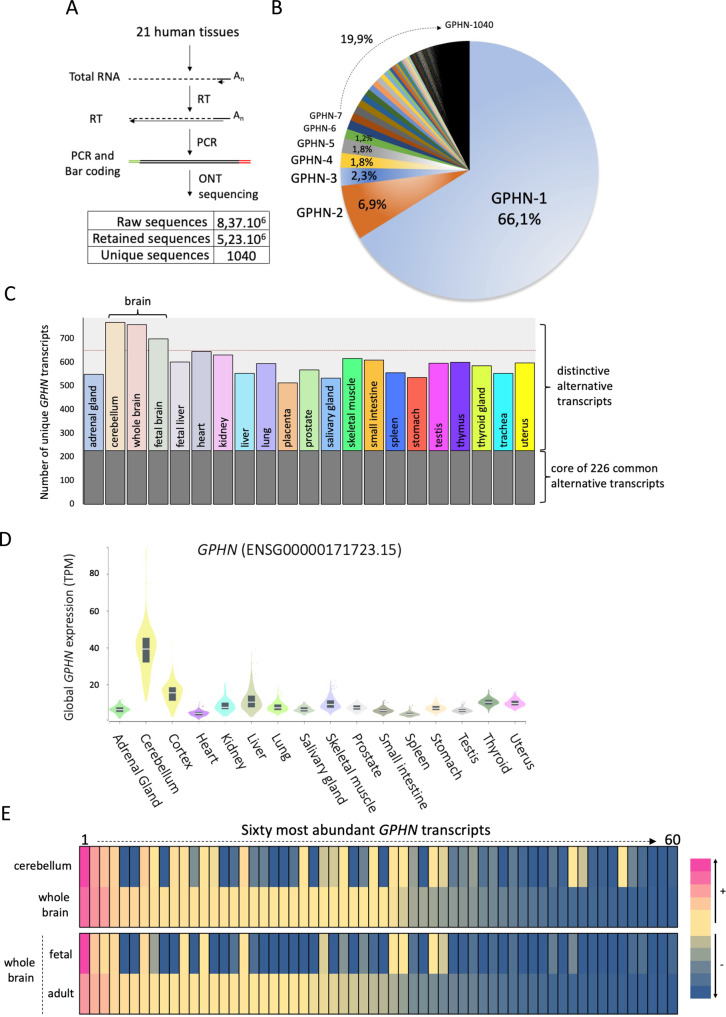
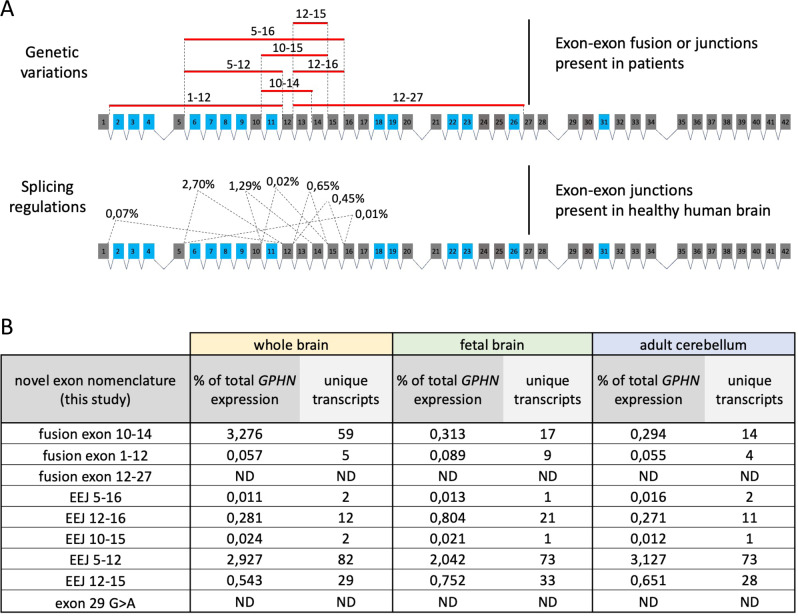
Gephyrin (GPHN) regulates the clustering of postsynaptic components at inhibitory synapses and is involved in pathophysiology of neuropsychiatric disorders. Here, we uncover an extensive diversity of GPHN transcripts that are tightly controlled by splicing during mouse and human brain development. Proteomic analysis reveals at least a hundred isoforms of GPHN incorporated at inhibitory Glycine and gamma-aminobutyric acid A receptors containing synapses. They exhibit different localization and postsynaptic clustering properties, and altering the expression level of one isoform is sufficient to affect the number, size, and density of inhibitory synapses in cerebellar Purkinje cells. Furthermore, we discovered that splicing defects reported in neuropsychiatric disorders are carried by multiple alternative GPHN transcripts, demonstrating the need for a thorough analysis of the GPHN transcriptome in patients. Overall, we show that alternative splicing of GPHN is an important genetic variation to consider in neurological diseases and a determinant of the diversity of postsynaptic inhibitory synapses.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Automated Image Analysis Reveals Different Localization of Synaptic Gephyrin C4 Splice Variants.eNeuro. 2023 Jan 4;10(1):ENEURO.0102-22.2022. doi: 10.1523/ENEURO.0102-22.2022. Print 2023 Jan. eNeuro. 2023. PMID: 36543537 Free PMC article.
-
Isoform heterogeneity of the human gephyrin gene (GPHN), binding domains to the glycine receptor, and mutation analysis in hyperekplexia.J Biol Chem. 2003 Jul 4;278(27):24688-96. doi: 10.1074/jbc.M301070200. Epub 2003 Apr 8. J Biol Chem. 2003. PMID: 12684523
-
Glycine and GABA(A) receptor subunits on Renshaw cells: relationship with presynaptic neurotransmitters and postsynaptic gephyrin clusters.J Comp Neurol. 2002 Mar 12;444(3):275-89. doi: 10.1002/cne.10148. J Comp Neurol. 2002. PMID: 11840480
-
Gephyrin and the regulation of synaptic strength and dynamics at glycinergic inhibitory synapses.Brain Res Bull. 2017 Mar;129:50-65. doi: 10.1016/j.brainresbull.2016.09.003. Epub 2016 Sep 6. Brain Res Bull. 2017. PMID: 27612963 Review.
-
Gephyrin: a master regulator of neuronal function?Nat Rev Neurosci. 2014 Mar;15(3):141-56. doi: 10.1038/nrn3670. Nat Rev Neurosci. 2014. PMID: 24552784 Review.
Cited by
-
Automated Image Analysis Reveals Different Localization of Synaptic Gephyrin C4 Splice Variants.eNeuro. 2023 Jan 4;10(1):ENEURO.0102-22.2022. doi: 10.1523/ENEURO.0102-22.2022. Print 2023 Jan. eNeuro. 2023. PMID: 36543537 Free PMC article.
-
Neurobeachin regulates receptor downscaling at GABAergic inhibitory synapses in a protein kinase A-dependent manner.Commun Biol. 2024 Dec 12;7(1):1635. doi: 10.1038/s42003-024-07294-z. Commun Biol. 2024. PMID: 39668217 Free PMC article.
-
Functional Neuroligin-2-MDGA1 interactions differentially regulate synaptic GABAARs and cytosolic gephyrin aggregation.Commun Biol. 2024 Sep 17;7(1):1157. doi: 10.1038/s42003-024-06789-z. Commun Biol. 2024. PMID: 39284869 Free PMC article.
-
Gephyrin promotes autonomous assembly and synaptic localization of GABAergic postsynaptic components without presynaptic GABA release.Proc Natl Acad Sci U S A. 2024 Jun 25;121(26):e2315100121. doi: 10.1073/pnas.2315100121. Epub 2024 Jun 18. Proc Natl Acad Sci U S A. 2024. PMID: 38889143 Free PMC article.
-
Revisiting the development of cerebellar inhibitory interneurons in the light of single-cell genetic analyses.Histochem Cell Biol. 2024 Jan;161(1):5-27. doi: 10.1007/s00418-023-02251-z. Epub 2023 Nov 8. Histochem Cell Biol. 2024. PMID: 37940705 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
