

A combined human gastruloid model of cardiogenesis and neurogenesis
- PMID: 35721464
- PMCID: PMC9198845
- DOI: 10.1016/j.isci.2022.104486
A combined human gastruloid model of cardiogenesis and neurogenesis
Abstract
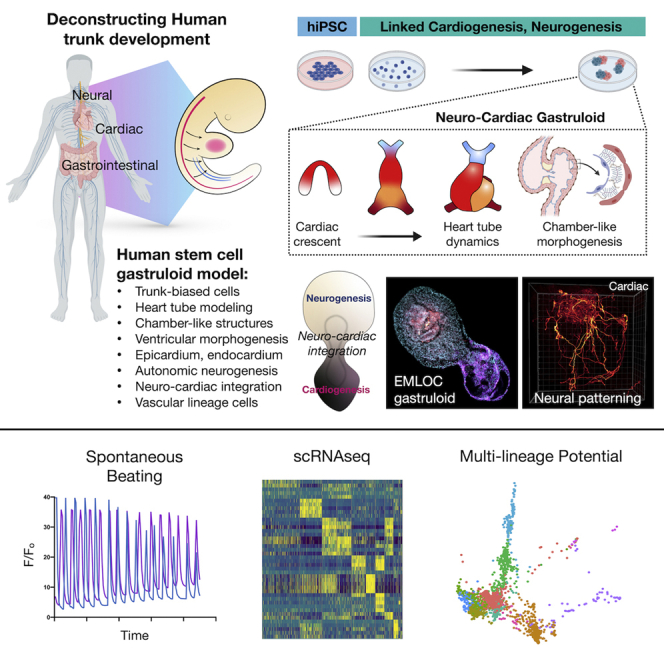
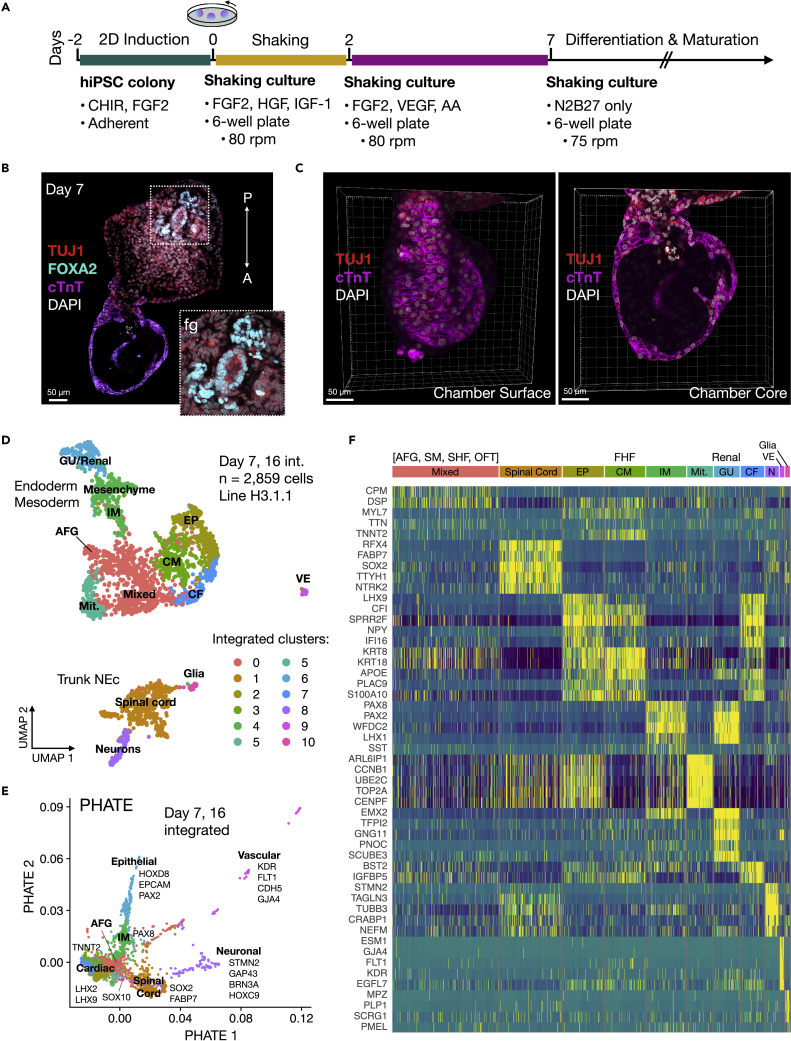
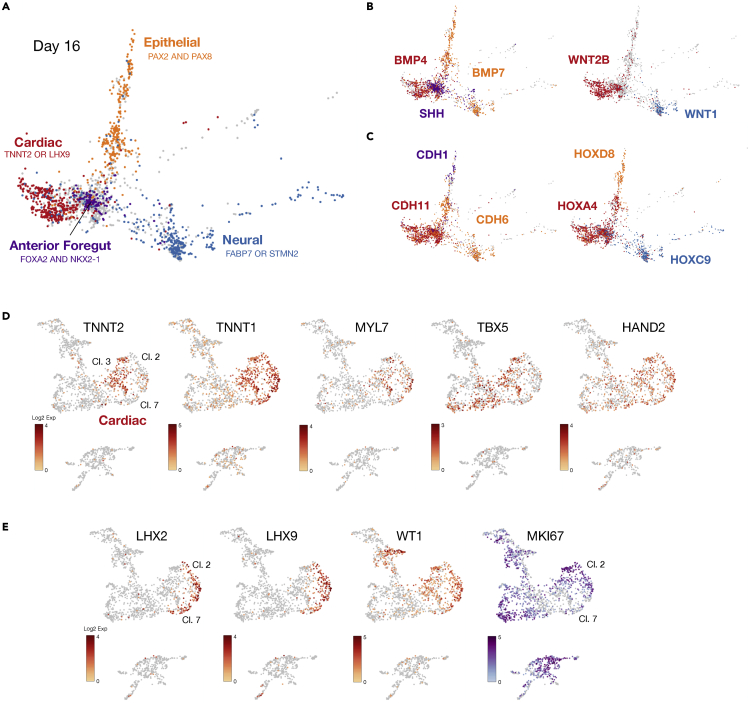
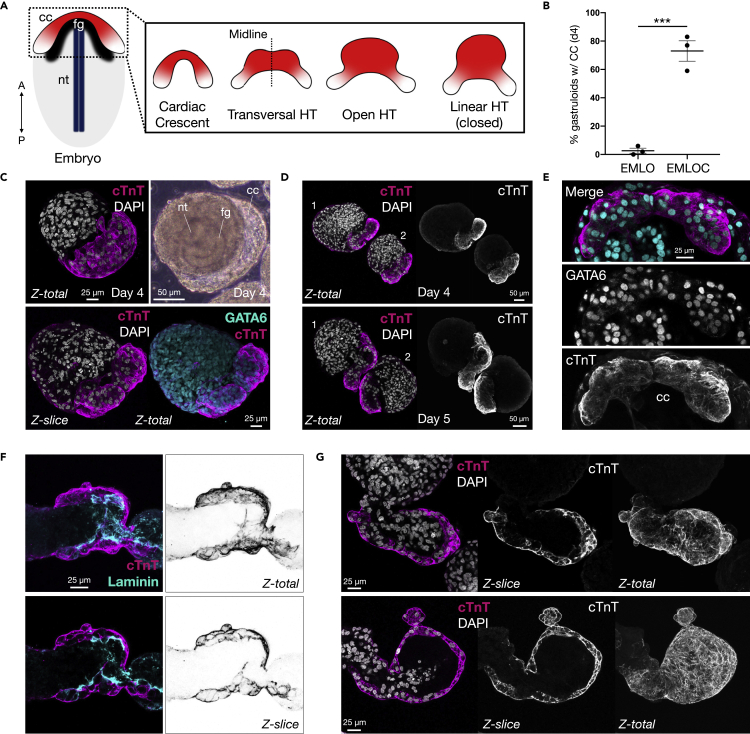
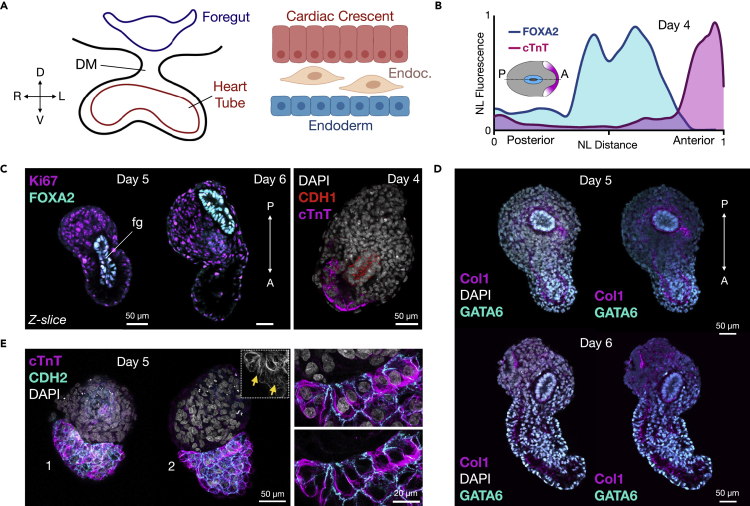
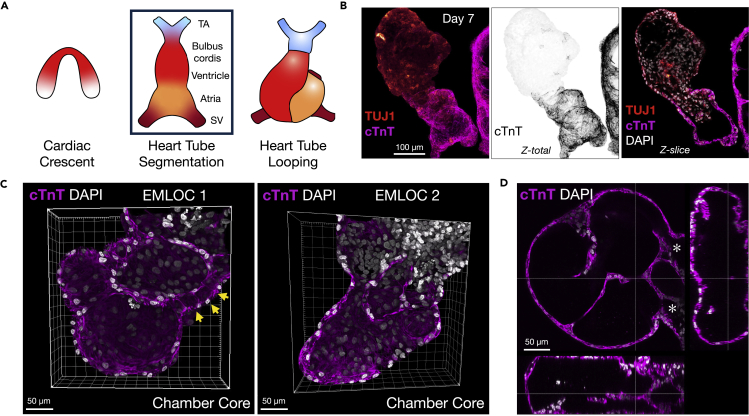
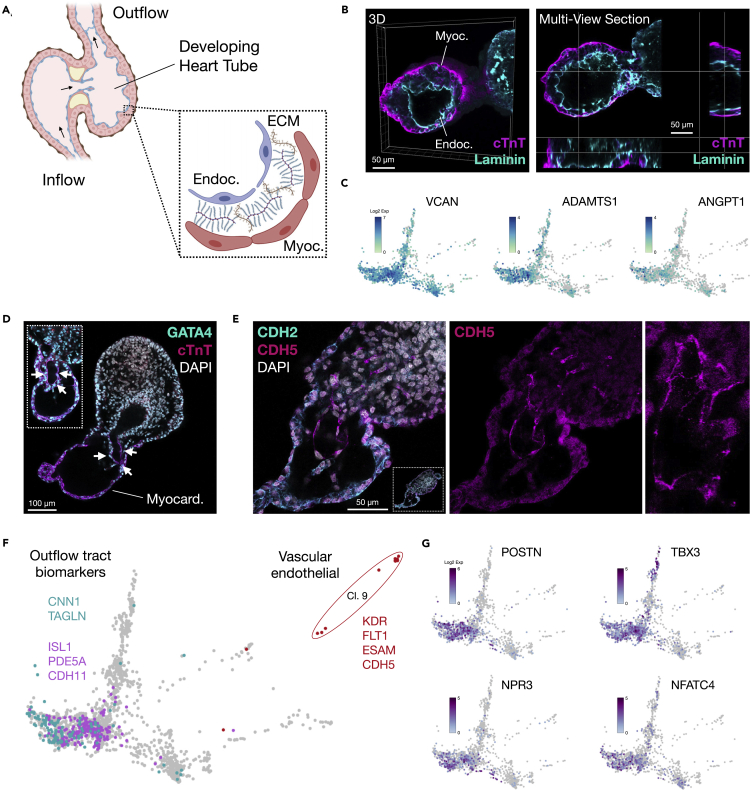
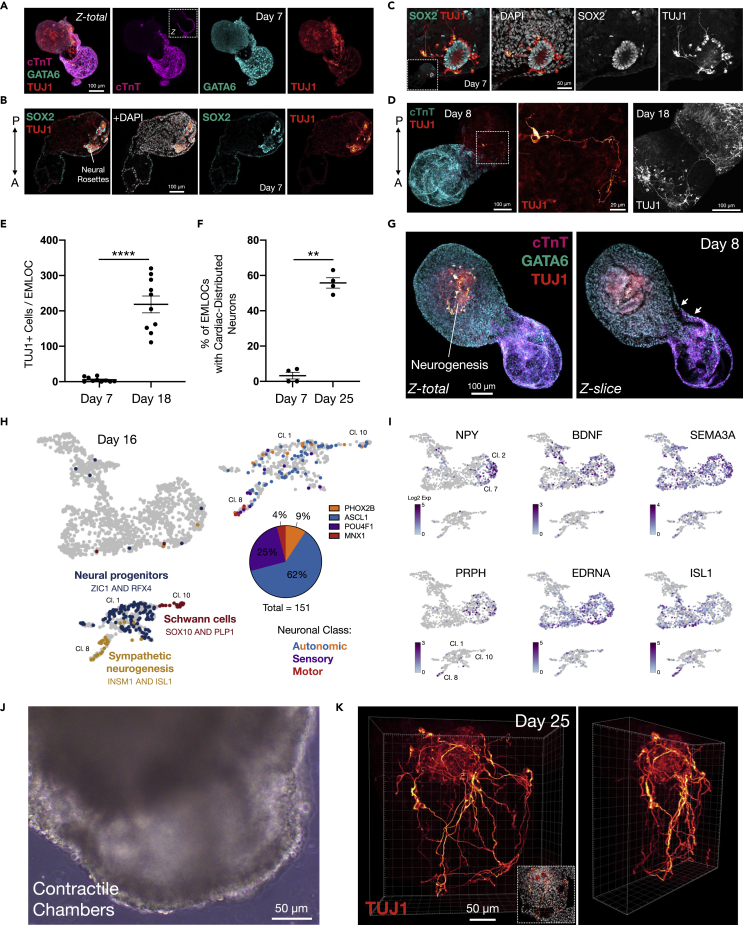
Multi-lineage development from gastruloids is enabling unprecedented opportunities to model and study human embryonic processes and is expected to accelerate ex vivo strategies in organ development. Reproducing human cardiogenesis with neurogenesis in a multi-lineage context remains challenging, requiring spatiotemporal input of paracrine and mechanical cues. Here we extend elongating multi-lineage organized (EMLO) gastruloids to include cardiogenesis (EMLOC) and describe interconnected neuro-cardiac lineages in a single gastruloid model. Contractile EMLOCs recapitulate numerous interlinked developmental features including heart tube formation and specialization, cardiomyocyte differentiation and remodeling phases, epicardium, ventricular wall morphogenesis, chamber-like structures and formation of a putative outflow tract. The EMLOC cardiac region, which originates anterior to gut tube primordium, is progressively populated by neurons in a spatial pattern mirroring the known distribution of neurons in the innervated human heart. This human EMLOC model represents a multi-lineage advancement for the study of coincident neurogenesis and cardiogenesis.
Keywords: Cell biology; Genomics; Stem cells research.
© 2022 The Author(s).
Conflict of interest statement
All affiliations are listed on the title page of the manuscript. All funding sources for this study are listed in the “acknowledgments” section of the manuscript. We, the authors and our immediate family members, have no financial intersts to declare. We, the authors and our immediate family members, have no positions to declare and are not members of the journal’s advisory board. A provisional USPTO patent on EMLOCs has been filed with patent application number 63/311,498. The authors declare no other competing interests.
Figures
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
