

Identification of Valerate as Carrying Capacity Modulator by Analyzing Lactiplantibacillus plantarum Colonization of Colonic Microbiota in vitro
- PMID: 35722334
- PMCID: PMC9197689
- DOI: 10.3389/fmicb.2022.910609
Identification of Valerate as Carrying Capacity Modulator by Analyzing Lactiplantibacillus plantarum Colonization of Colonic Microbiota in vitro
Abstract
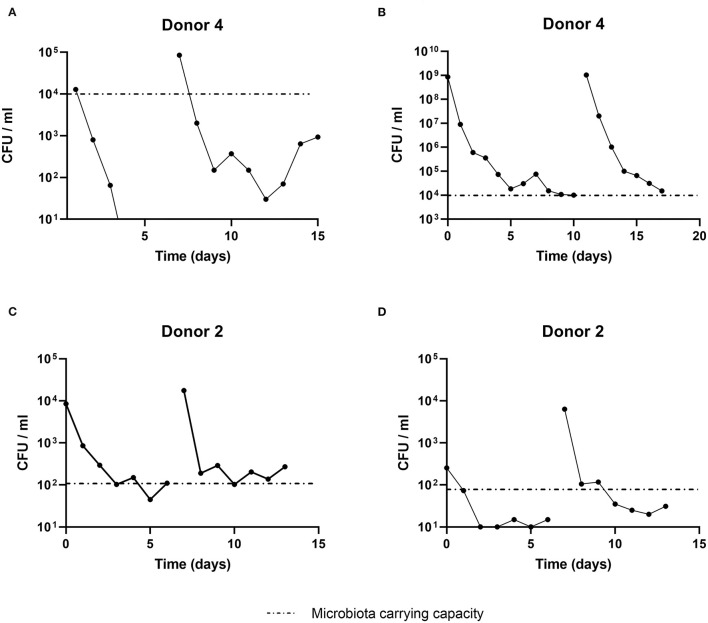
Humans ingest many microorganisms, which may colonize and interact with the resident gut microbiota. However, extensive knowledge about host-independent microbe-microbe interactions is lacking. Here, we investigated such colonization process using a derivative of the model probiotic Lactiplantibacillus plantarum WCFS1 into continuously cultivated gut microbiota in the intestinal PolyFermS fermentation model inoculated with five independently immobilized human adult fecal microbiota. L. plantarum successfully colonized and organized itself spatially in the planktonic, that is, the reactor effluent, and sessile, that is, reactor biofilm, fractions of distinct human adult microbiota. The microbiota carrying capacity for L. plantarum was independent of L. plantarum introduction dose and second supplementation. Adult microbiota (n = 3) dominated by Prevotella and Ruminoccocus exhibited a higher carrying capacity than microbiota (n = 2) dominated by Bacteroides with 105 and 103 CFU/ml of L. plantarum, respectively. Cultivation of human adult microbiota over 3 months resulted in decreased carrying capacity and correlated positively with richness and evenness, suggesting enhanced resistance toward colonizers. Our analyses ultimately allowed us to identify the fermentation metabolite valerate as a modulator to increase the carrying capacity in a microbiota-independent manner. In conclusion, by uncoupling microbe-microbe interactions from host factors, we showed that L. plantarum colonizes the in vitro colonic community in a microbiota-dependent manner. We were further able to demonstrate that L. plantarum colonization levels were not susceptible to the introduction parameters dose and repeated administration but to microbiota features. Such knowledge is relevant in gaining a deeper ecological understanding of colonizer-microbiota interactions and developing robust probiotic strategies.
Keywords: PolyFermS; gut microbiome; intestinal ecology; microbiota invasion; probiotics.
Copyright © 2022 Isenring, Stevens, Jans, Lacroix and Geirnaert.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures





References
LinkOut - more resources
Full Text Sources