

Minimal functional domains of the core polarity regulator Dlg
- PMID: 35722710
- PMCID: PMC9346270
- DOI: 10.1242/bio.059408
Minimal functional domains of the core polarity regulator Dlg
Abstract
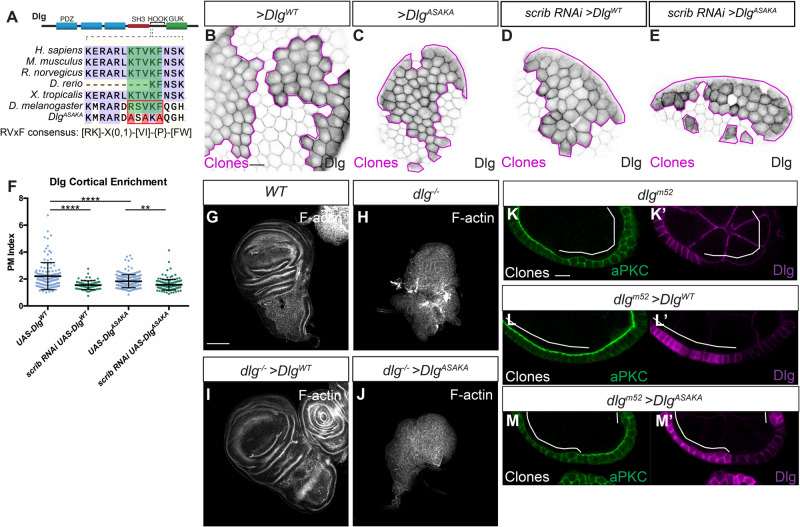
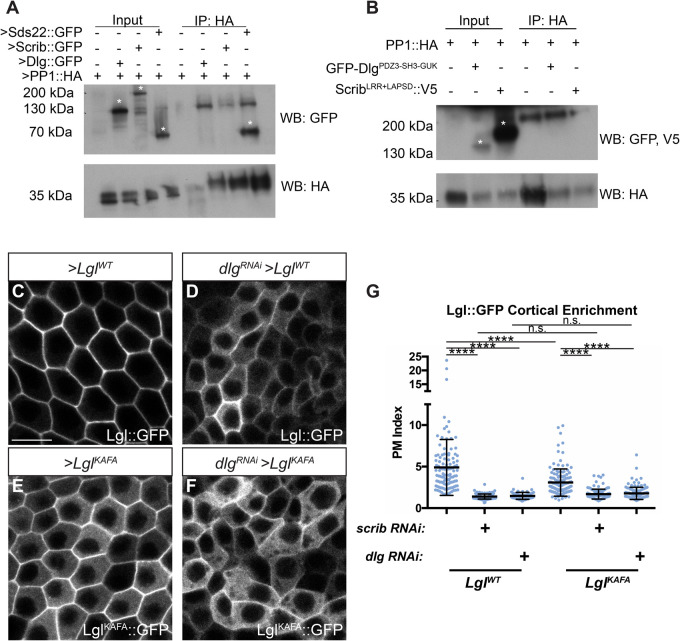
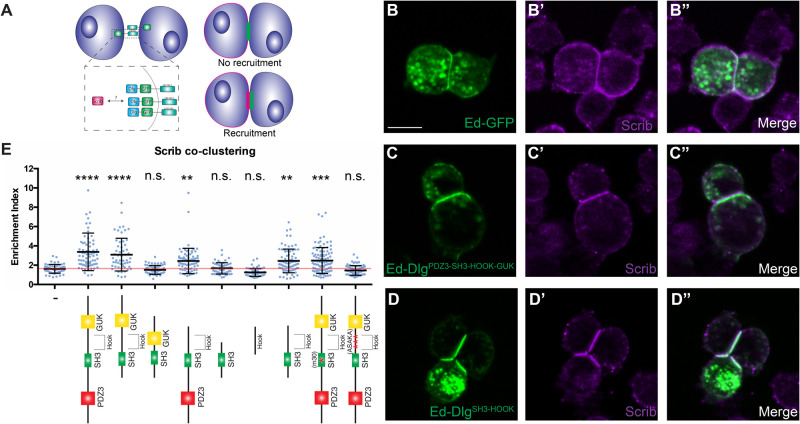
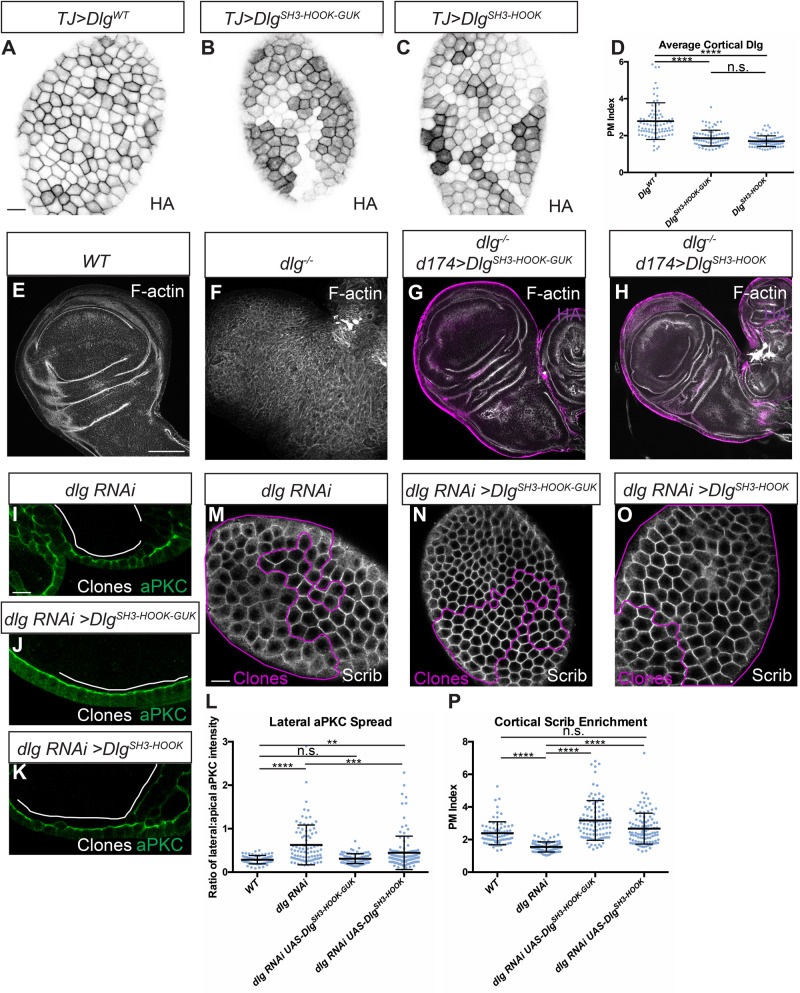
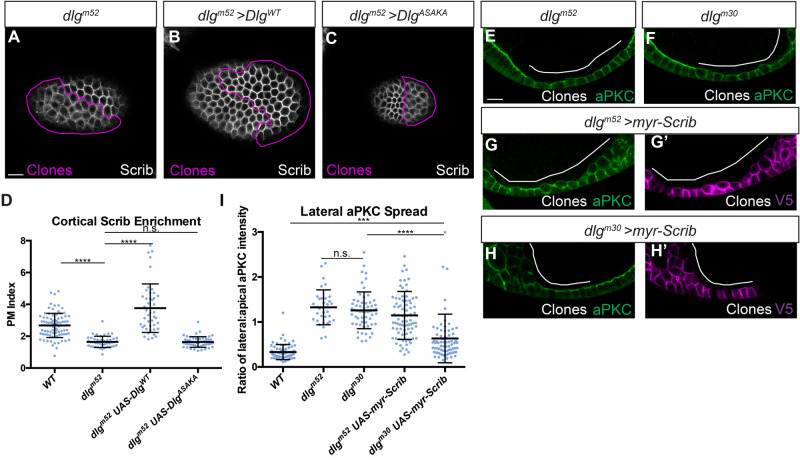
The compartmentalized domains of polarized epithelial cells arise from mutually antagonistic actions between the apical Par complex and the basolateral Scrib module. In Drosophila, the Scrib module proteins Scribble (Scrib) and Discs-large (Dlg) are required to limit Lgl phosphorylation at the basolateral cortex, but how Scrib and Dlg could carry out such a 'protection' activity is not clear. We tested Protein Phosphatase 1α (PP1) as a potential mediator of this activity, but demonstrate that a significant component of Scrib and Dlg regulation of Lgl is PP1 independent, and found no evidence for a Scrib-Dlg-PP1 protein complex. However, the Dlg SH3 domain plays a role in Lgl protection and, in combination with the N-terminal region of the Dlg HOOK domain, in recruitment of Scrib to the membrane. We identify a 'minimal Dlg' comprised of the SH3 and HOOK domains that is both necessary and sufficient for Scrib localization and epithelial polarity function in vivo. This article has an associated First Person interview with the first author of the paper.
Keywords: Cell; Dlg; Polarity; Scrib; Scrib module.
© 2022. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interests The authors declare no competing or financial interests.
Figures
Similar articles
-
Lgl cortical dynamics are independent of binding to the Scrib-Dlg complex but require Dlg-dependent restriction of aPKC.Development. 2020 Aug 7;147(15):dev186593. doi: 10.1242/dev.186593. Development. 2020. PMID: 32665243
-
Distinct activities of Scrib module proteins organize epithelial polarity.Proc Natl Acad Sci U S A. 2020 May 26;117(21):11531-11540. doi: 10.1073/pnas.1918462117. Epub 2020 May 15. Proc Natl Acad Sci U S A. 2020. PMID: 32414916 Free PMC article.
-
Drosophila melanogaster Guk-holder interacts with the Scribbled PDZ1 domain and regulates epithelial development with Scribbled and Discs Large.J Biol Chem. 2018 Mar 23;293(12):4519-4531. doi: 10.1074/jbc.M117.817528. Epub 2018 Jan 29. J Biol Chem. 2018. PMID: 29378849 Free PMC article.
-
Dlg, Scribble and Lgl in cell polarity, cell proliferation and cancer.Bioessays. 2003 Jun;25(6):542-53. doi: 10.1002/bies.10286. Bioessays. 2003. PMID: 12766944 Review.
-
Scrib module proteins: Control of epithelial architecture and planar spindle orientation.Int J Biochem Cell Biol. 2021 Jul;136:106001. doi: 10.1016/j.biocel.2021.106001. Epub 2021 May 4. Int J Biochem Cell Biol. 2021. PMID: 33962021 Review.
Cited by
-
A conditional strategy for cell-type-specific labeling of endogenous excitatory synapses in Drosophila.Cell Rep Methods. 2023 May 11;3(5):100477. doi: 10.1016/j.crmeth.2023.100477. eCollection 2023 May 22. Cell Rep Methods. 2023. PMID: 37323572 Free PMC article.
References
-
- Awadia, S., Huq, F., Arnold, T. R., Goicoechea, S. M., Sun, Y. J., Hou, T., Kreider-Letterman, G., Massimi, P., Banks, L., Fuentes, E. J.et al. (2019). SGEF forms a complex with Scribble and Dlg1 and regulates epithelial junctions and contractility. J. Cell Biol. 218, 2699-2725. 10.1083/jcb.201811114 - DOI - PMC - PubMed
-
- Bellaïche, Y., Radovic, A., Woods, D. F., Hough, C. D., Parmentier, M.-L., O'Kane, C. J., Bryant, P. J. and Schweisguth, F. (2001). The partner of inscuteable/discs-large complex is required to establish planar polarity during asymmetric cell division in Drosophila. Cell 106, 355-366. 10.1016/S0092-8674(01)00444-5 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
