

Missense variants causing Wiedemann-Steiner syndrome preferentially occur in the KMT2A-CXXC domain and are accurately classified using AlphaFold2
- PMID: 35727845
- PMCID: PMC9249231
- DOI: 10.1371/journal.pgen.1010278
Missense variants causing Wiedemann-Steiner syndrome preferentially occur in the KMT2A-CXXC domain and are accurately classified using AlphaFold2
Abstract
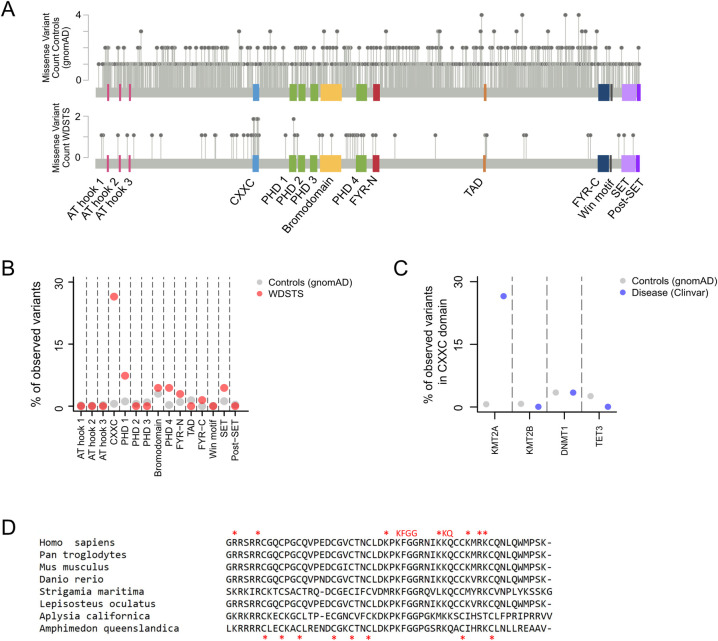
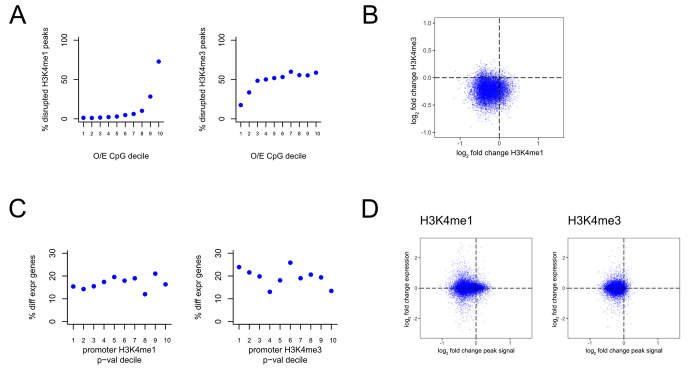
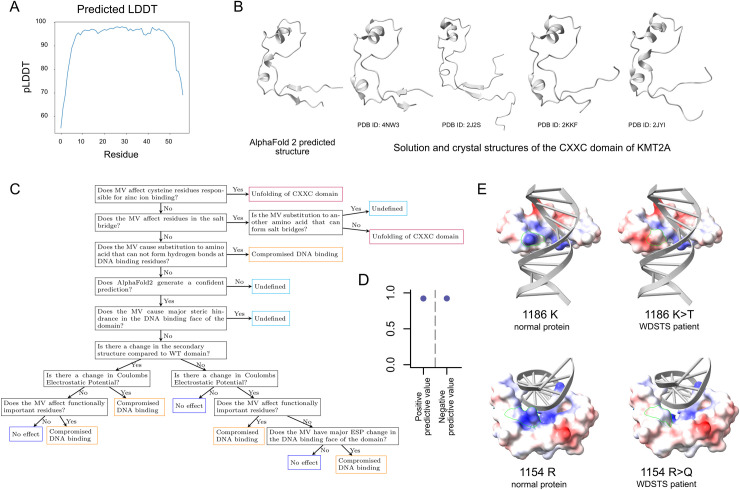
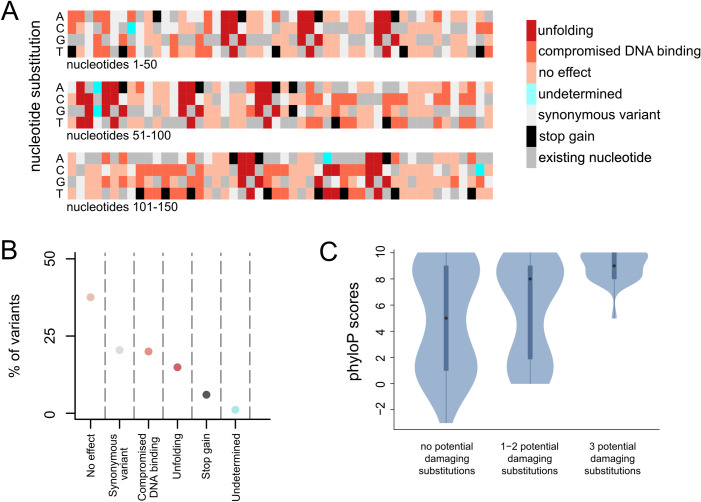
Wiedemann-Steiner syndrome (WDSTS) is a neurodevelopmental disorder caused by de novo variants in KMT2A, which encodes a multi-domain histone methyltransferase. To gain insight into the currently unknown pathogenesis of WDSTS, we examined the spatial distribution of likely WDSTS-causing variants across the 15 different domains of KMT2A. Compared to variants in healthy controls, WDSTS variants exhibit a 61.9-fold overrepresentation within the CXXC domain-which mediates binding to unmethylated CpGs-suggesting a major role for this domain in mediating the phenotype. In contrast, we find no significant overrepresentation within the catalytic SET domain. Corroborating these results, we find that hippocampal neurons from Kmt2a-deficient mice demonstrate disrupted histone methylation (H3K4me1 and H3K4me3) preferentially at CpG-rich regions, but this has no systematic impact on gene expression. Motivated by these results, we combine accurate prediction of the CXXC domain structure by AlphaFold2 with prior biological knowledge to develop a classification scheme for missense variants in the CXXC domain. Our classifier achieved 92.6% positive and 92.9% negative predictive value on a hold-out test set. This classification performance enabled us to subsequently perform an in silico saturation mutagenesis and classify a total of 445 variants according to their functional effects. Our results yield a novel insight into the mechanistic basis of WDSTS and provide an example of how AlphaFold2 can contribute to the in silico characterization of variant effects with very high accuracy, suggesting a paradigm potentially applicable to many other Mendelian disorders.
Conflict of interest statement
I have read the journal’s policy and the authors of this manuscript have the following competing interests: HTB is a consultant for Mahzi therapeutics. No other authors have any potential conflict of interest.
Figures
References
-
- Voo K.S., et al.., Cloning of a mammalian transcriptional activator that binds unmethylated CpG motifs and shares a CXXC domain with DNA methyltransferase, human trithorax, and methyl-CpG binding domain protein 1. Mol Cell Biol, 2000. 20(6): p. 2108–21. doi: 10.1128/MCB.20.6.2108-2121.2000 - DOI - PMC - PubMed
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
