

Dietary folate drives methionine metabolism to promote cancer development by stabilizing MAT IIA
- PMID: 35729157
- PMCID: PMC9213445
- DOI: 10.1038/s41392-022-01017-8
Dietary folate drives methionine metabolism to promote cancer development by stabilizing MAT IIA
Erratum in
-
Correction To: Dietary folate drives methionine metabolism to promote cancer development by stabilizing MAT IIA.Signal Transduct Target Ther. 2022 Dec 28;7(1):401. doi: 10.1038/s41392-022-01255-w. Signal Transduct Target Ther. 2022. PMID: 36577756 Free PMC article. No abstract available.
Abstract
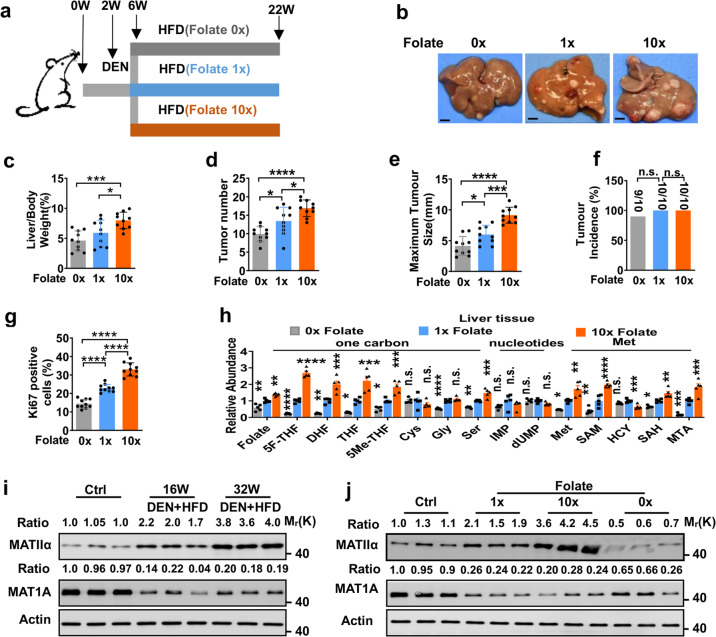
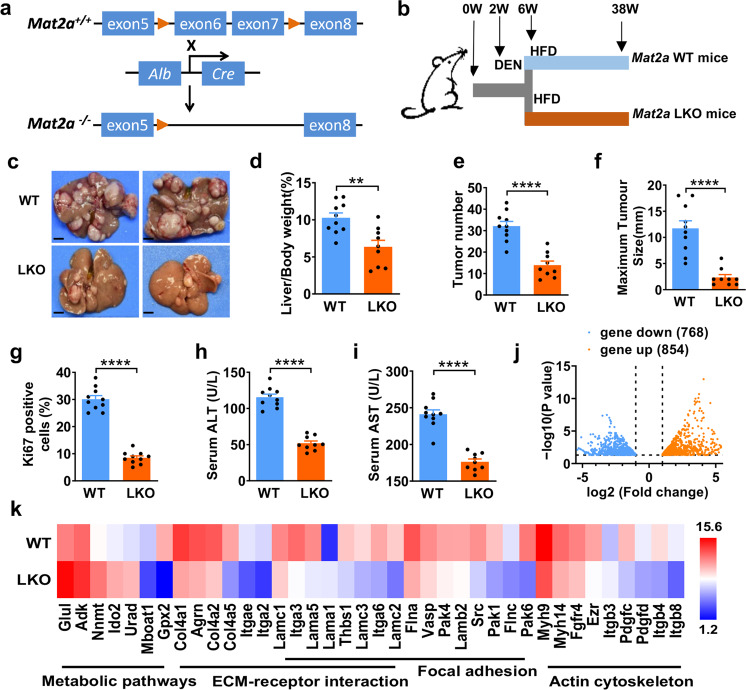
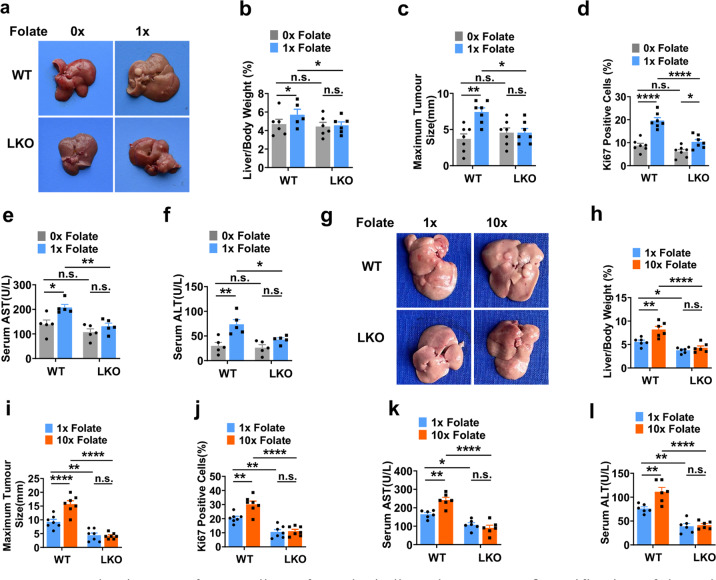
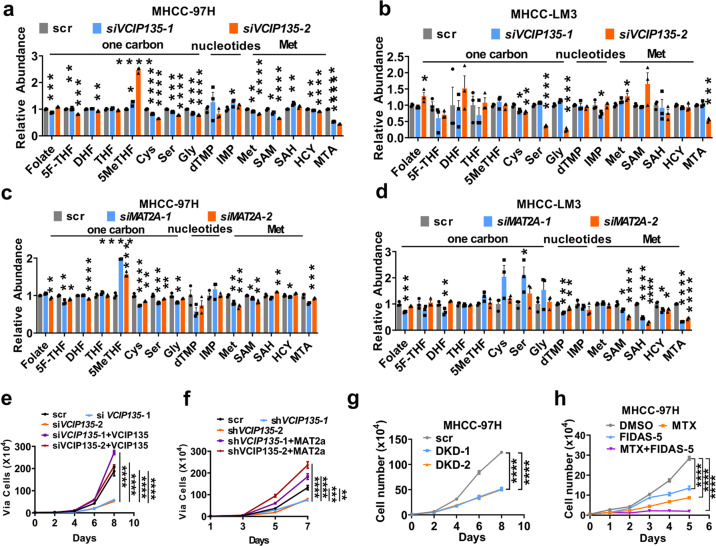
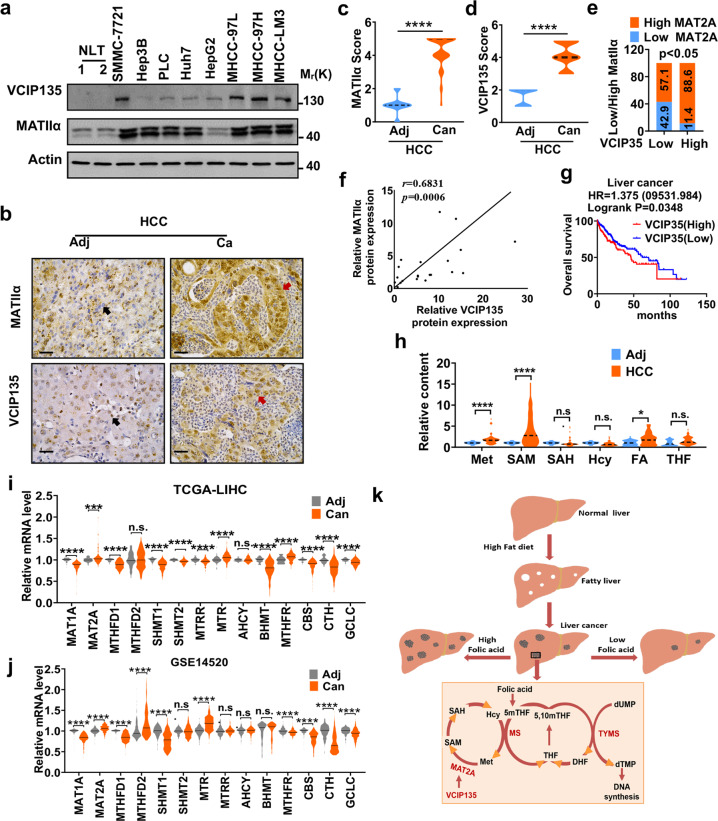
Folic acid, served as dietary supplement, is closely linked to one-carbon metabolism and methionine metabolism. Previous clinical evidence indicated that folic acid supplementation displays dual effect on cancer development, promoting or suppressing tumor formation and progression. However, the underlying mechanism remains to be uncovered. Here, we report that high-folate diet significantly promotes cancer development in mice with hepatocellular carcinoma (HCC) induced by DEN/high-fat diet (HFD), simultaneously with increased expression of methionine adenosyltransferase 2A (gene name, MAT2A; protein name, MATIIα), the key enzyme in methionine metabolism, and acceleration of methionine cycle in cancer tissues. In contrast, folate-free diet reduces MATIIα expression and impedes HFD-induced HCC development. Notably, methionine metabolism is dynamically reprogrammed with valosin-containing protein p97/p47 complex-interacting protein (VCIP135) which functions as a deubiquitylating enzyme to bind and stabilize MATIIα in response to folic acid signal. Consistently, upregulation of MATIIα expression is positively correlated with increased VCIP135 protein level in human HCC tissues compared to adjacent tissues. Furthermore, liver-specific knockout of Mat2a remarkably abolishes the advocating effect of folic acid on HFD-induced HCC, demonstrating that the effect of high or free folate-diet on HFD-induced HCC relies on Mat2a. Moreover, folate and multiple intermediate metabolites in one-carbon metabolism are significantly decreased in vivo and in vitro upon Mat2a deletion. Together, folate promotes the integration of methionine and one-carbon metabolism, contributing to HCC development via hijacking MATIIα metabolic pathway. This study provides insight into folate-promoted cancer development, strongly recommending the tailor-made folate supplement guideline for both sub-healthy populations and patients with cancer expressing high level of MATIIα expression.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests. Q.-Y.L. is the editorial board member of Signal Transduction and Targeted Therapy, but she has not been involved in the process of the manuscript handling.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
