

Neutralizing Antibodies against Lassa Virus Lineage I
- PMID: 35730904
- PMCID: PMC9426445
- DOI: 10.1128/mbio.01278-22
Neutralizing Antibodies against Lassa Virus Lineage I
Abstract
Lassa virus (LASV) is the causative agent of the deadly Lassa fever (LF). Seven distinct LASV lineages circulate through western Africa, among which lineage I (LI), the first to be identified, is particularly resistant to antibody neutralization. Lineage I LASV evades neutralization by half of known antibodies in the GPC-A antibody competition group and all but one of the antibodies in the GPC-B competition group. Here, we solve two cryo-electron microscopy (cryo-EM) structures of LI GP in complex with a GPC-A and a GPC-B antibody. We used complementary structural and biochemical techniques to identify single-amino-acid substitutions in LI that are responsible for immune evasion by each antibody group. Further, we show that LI infection is more dependent on the endosomal receptor lysosome-associated membrane protein 1 (LAMP1) for viral entry relative to LIV. In the absence of LAMP1, LI requires a more acidic fusion pH to initiate membrane fusion with the host cell relative to LIV. IMPORTANCE No vaccine or therapeutics are approved to prevent LASV infection or treat LF. All vaccine platforms currently under development present only the LIV GP sequence. However, our data suggest that the high genetic diversity of LASV may be problematic for designing both a broadly reactive immunogen and therapeutic. Here, we examine antibodies that are highly potent against LIV yet are ineffective against LI. By pinpointing LI mutations responsible for this decrease in antibody efficacy, we suggest that future vaccine platforms may need to incorporate specific LI-like mutations in order to generate a broadly neutralizing antibody response against all LASV lineages.
Keywords: LAMP1; Lassa fever; Lassa virus; antigenic variation; cryo-EM; hemorrhagic fever virus; neutralizing antibodies; prefusion glycoprotein; structure-based vaccine design; structure-guided immunogen; viral escape.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
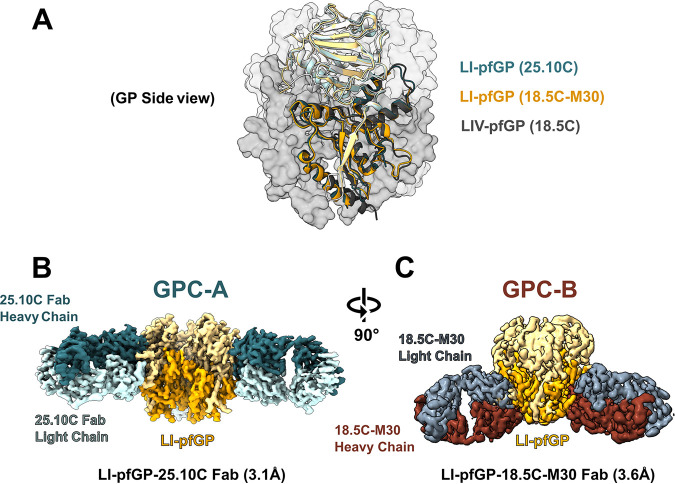
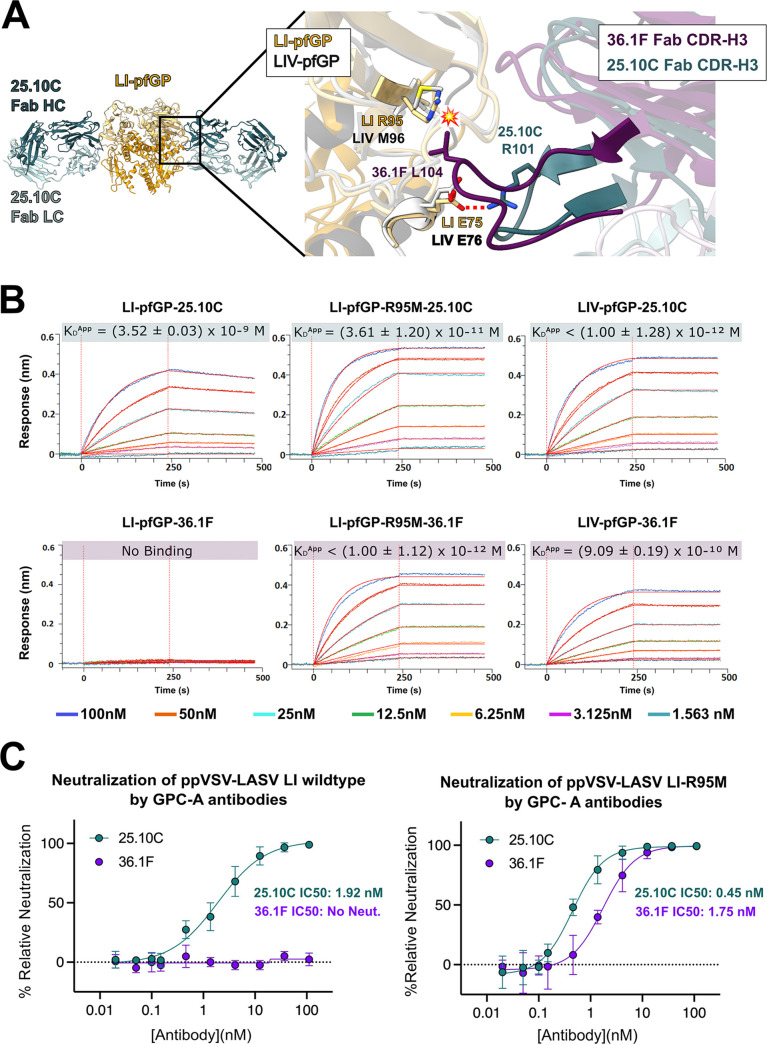
Similar articles
-
Delineating the mechanism of anti-Lassa virus GPC-A neutralizing antibodies.Cell Rep. 2022 May 24;39(8):110841. doi: 10.1016/j.celrep.2022.110841. Cell Rep. 2022. PMID: 35613585 Free PMC article.
-
Structural conservation of Lassa virus glycoproteins and recognition by neutralizing antibodies.Cell Rep. 2023 May 30;42(5):112524. doi: 10.1016/j.celrep.2023.112524. Epub 2023 May 18. Cell Rep. 2023. PMID: 37209096 Free PMC article.
-
Lamp1 Increases the Efficiency of Lassa Virus Infection by Promoting Fusion in Less Acidic Endosomal Compartments.mBio. 2018 Jan 2;9(1):e01818-17. doi: 10.1128/mBio.01818-17. mBio. 2018. PMID: 29295909 Free PMC article.
-
Inter-Lineage Variation of Lassa Virus Glycoprotein Epitopes: A Challenge to Lassa Virus Vaccine Development.Viruses. 2020 Mar 31;12(4):386. doi: 10.3390/v12040386. Viruses. 2020. PMID: 32244402 Free PMC article. Review.
-
Infusion of neutralization into Lassa vaccine design.Trends Immunol. 2025 Jul;46(7):512-524. doi: 10.1016/j.it.2025.05.006. Epub 2025 Jun 24. Trends Immunol. 2025. PMID: 40555565 Review.
Cited by
-
Advancements and future prospects of adeno-associated virus-mediated gene therapy for sensorineural hearing loss.Front Neurosci. 2024 Jan 24;18:1272786. doi: 10.3389/fnins.2024.1272786. eCollection 2024. Front Neurosci. 2024. PMID: 38327848 Free PMC article. Review.
-
Mapping the antibody response to Lassa virus vaccination of non-human primates.EBioMedicine. 2025 Apr;114:105673. doi: 10.1016/j.ebiom.2025.105673. Epub 2025 Mar 31. EBioMedicine. 2025. PMID: 40168843 Free PMC article.
-
Molecular organization of the New World arenavirus spike glycoprotein complex.Nat Microbiol. 2025 Sep;10(9):2207-2220. doi: 10.1038/s41564-025-02085-6. Epub 2025 Aug 8. Nat Microbiol. 2025. PMID: 40781447 Free PMC article.
-
Deep mutational scanning reveals functional constraints and antibody-escape potential of Lassa virus glycoprotein complex.Immunity. 2024 Sep 10;57(9):2061-2076.e11. doi: 10.1016/j.immuni.2024.06.013. Epub 2024 Jul 15. Immunity. 2024. PMID: 39013466 Free PMC article.
-
Entry inhibitors as arenavirus antivirals.Front Microbiol. 2024 Apr 8;15:1382953. doi: 10.3389/fmicb.2024.1382953. eCollection 2024. Front Microbiol. 2024. PMID: 38650890 Free PMC article. Review.
References
-
- Shaffer JG, Grant DS, Schieffelin JS, Boisen ML, Goba A, Hartnett JN, Levy DC, Yenni RE, Moses LM, Fullah M, Momoh M, Fonnie M, Fonnie R, Kanneh L, Koroma VJ, Kargbo K, Ottomassathien D, Muncy IJ, Jones AB, Illick MM, Kulakosky PC, Haislip AM, Bishop CM, Elliot DH, Brown BL, Zhu H, Hastie KM, Andersen KG, Gire SK, Tabrizi S, Tariyal R, Stremlau M, Matschiner A, Sampey DB, Spence JS, Cross RW, Geisbert JB, Folarin OA, Happi CT, Pitts KR, Geske FJ, Geisbert TW, Saphire EO, Robinson JE, Wilson RB, Sabeti PC, Henderson LA, Khan SH, Bausch DG, Branco LM, Garry RF, Viral Hemorrhagic Fever Consortium . 2014. Lassa fever in post-conflict Sierra Leone. PLoS Negl Trop Dis 8:e2748. doi:10.1371/journal.pntd.0002748. - DOI - PMC - PubMed
-
- Ehichioya DU, Dellicour S, Pahlmann M, Rieger T, Oestereich L, Becker-Ziaja B, Cadar D, Ighodalo Y, Olokor T, Omomoh E, Oyakhilome J, Omiunu R, Agbukor J, Ebo B, Aiyepada J, Ebhodaghe P, Osiemi B, Ehikhametalor S, Akhilomen P, Airende M, Esumeh R, Muoebonam E, Giwa R, Ekanem A, Igenegbale G, Odigie G, Okonofua G, Enigbe R, Omonegho Yerumoh E, Pallasch E, Bockholt S, Kafetzopoulou LE, Duraffour S, Okokhere PO, Akpede GO, Okogbenin SA, Odia I, Aire C, Akpede N, Tobin E, Ogbaini-Emovon E, Lemey P, Adomeh DI, Asogun DA, Günther S. 2019. Phylogeography of Lassa virus in Nigeria. J Virol 93:e00929-19. doi:10.1128/JVI.00929-19. - DOI - PMC - PubMed
-
- Heinrich ML, Boisen ML, Nelson DKS, Bush DJ, Cross RW, Koval AP, Hoffmann AR, Beddingfield BJ, Hastie KM, Rowland MM, Aimukanova I, Koval S, Lathigra R, Borisevich V, Momoh M, Sandi JD, Goba A, Odia L, Baimba F, Aiyepada JO, Ebo B, Eromon P, Ugwu C, Folarin O, Olumade T, Onyechi MN, Etafo J, Adeyemi R, Ella EE, Aminu M, Gomerep SS, Eke MA, Ogunsanya O, Akpede GO, Asogun DO, Okogbenin SA, Okokhere PO, Holst J, Shaffer JG, Schieffelin JS, Geisbert TW, Saphire EO, Happi CT, Grant DS, Garry RF, Branco LM. 2020. Antibodies from Sierra Leonean and Nigerian Lassa fever survivors cross-react with recombinant proteins representing Lassa viruses of divergent lineages. Sci Rep 10:16030. doi:10.1038/s41598-020-72539-w. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
