

Complex floral traits shape pollinator attraction to ornamental plants
- PMID: 35732011
- PMCID: PMC9510942
- DOI: 10.1093/aob/mcac082
Complex floral traits shape pollinator attraction to ornamental plants
Abstract
Background and aims: Ornamental flowering plant species are often used in managed greenspaces to attract and support pollinator populations. In natural systems, selection by pollinators is hypothesized to result in convergent multimodal floral phenotypes that are more attractive to specific pollinator taxa. In contrast, ornamental cultivars are bred via artificial selection by humans, and exhibit diverse and distinct phenotypes. Despite their prevalence in managed habitats, the influence of cultivar phenotypic variation on plant attractiveness to pollinator taxa is not well resolved.
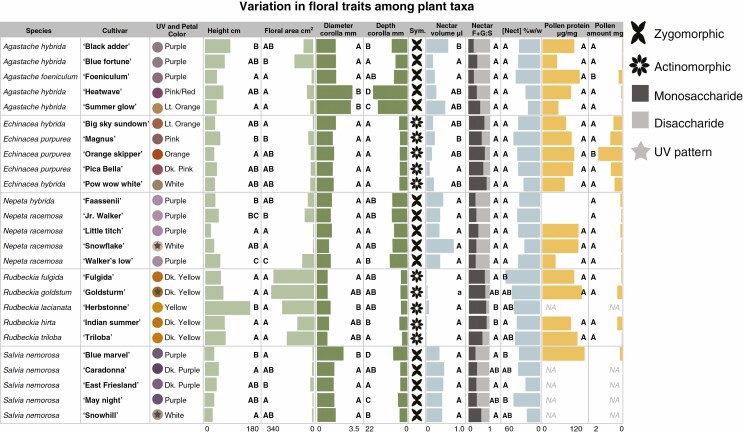
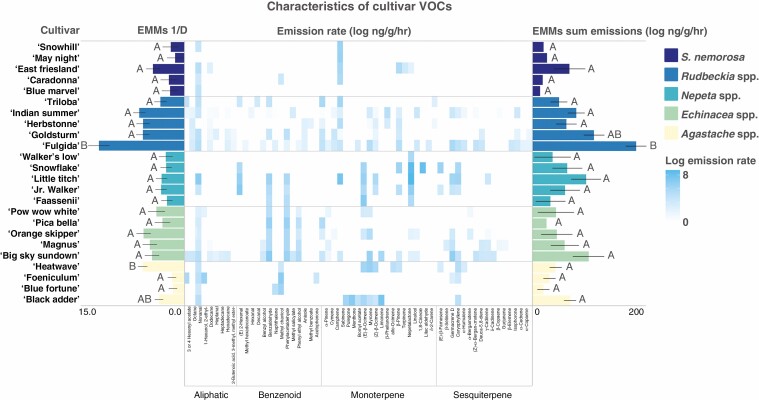
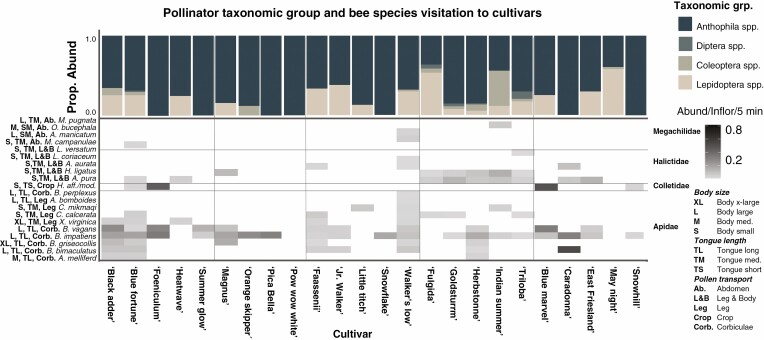
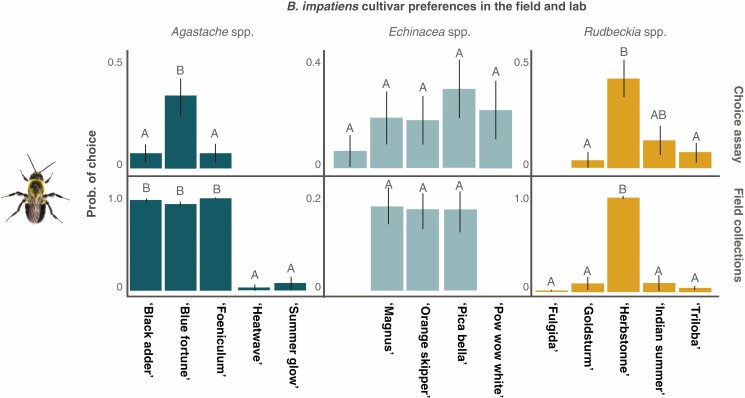
Methods: We used a combination of field and behavioural assays to evaluate how variation in floral visual, chemical and nutritional traits impacted overall attractiveness and visitation by pollinator taxonomic groups and bee species to 25 cultivars of five herbaceous perennial ornamental plant genera.
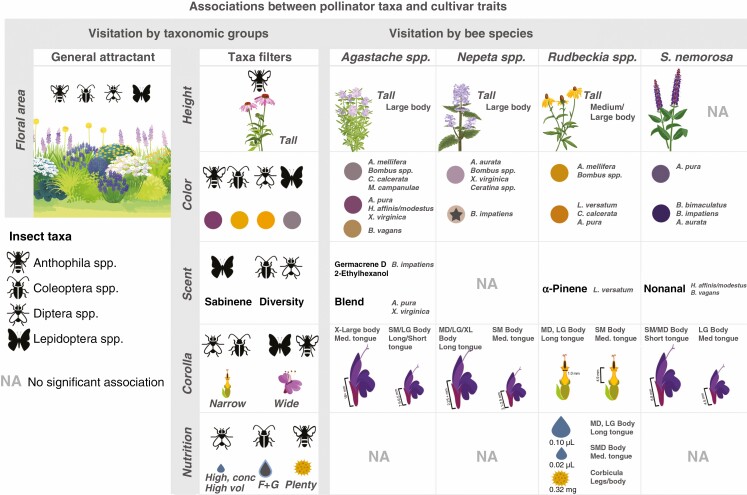
Key results: Despite significant phenotypic variation, cultivars tended to attract a broad range of pollinator species. Nonetheless, at the level of insect order (bee, fly, butterfly, beetle), attraction was generally modulated by traits consistent with the pollination syndrome hypothesis. At the level of bee species, the relative influence of traits on visitation varied across plant genera, with some floral phenotypes leading to a broadening of the visitor community, and others leading to exclusion of visitation by certain bee species.
Conclusions: Our results demonstrate how pollinator choice is mediated by complex multimodal floral signals. Importantly, the traits that had the greatest and most consistent effect on regulating pollinator attraction were those that are commonly selected for in cultivar development. Though variation among cultivars in floral traits may limit the pollinator community by excluding certain species, it may also encourage interactions with generalist taxa to support pollinator diversity in managed landscapes.
Keywords: Agastache spp; Echinacea spp; Nepeta spp; Rudbeckia spp; Salvia nemorosa; bees; floral traits; ornamentals; plant–pollinator interactions; pollination syndromes; pollinators.
© The Author(s) 2022. Published by Oxford University Press on behalf of the Annals of Botany Company.
Figures

Similar articles
-
Herbaceous perennial ornamental plants can support complex pollinator communities.Sci Rep. 2021 Aug 30;11(1):17352. doi: 10.1038/s41598-021-95892-w. Sci Rep. 2021. PMID: 34462447 Free PMC article.
-
More Than Meets the Eye? The Role of Annual Ornamental Flowers in Supporting Pollinators.Environ Entomol. 2020 Feb 17;49(1):178-188. doi: 10.1093/ee/nvz133. Environ Entomol. 2020. PMID: 31755522
-
Evolution and diversity of floral scent chemistry in the euglossine bee-pollinated orchid genus Gongora.Ann Bot. 2016 Jul;118(1):135-48. doi: 10.1093/aob/mcw072. Epub 2016 May 30. Ann Bot. 2016. PMID: 27240855 Free PMC article.
-
Why are there so many bee-orchid species? Adaptive radiation by intra-specific competition for mnesic pollinators.Biol Rev Camb Philos Soc. 2020 Dec;95(6):1630-1663. doi: 10.1111/brv.12633. Epub 2020 Sep 21. Biol Rev Camb Philos Soc. 2020. PMID: 32954662 Review.
-
Enhancing entomophilous pollination for sustainable crop production.Plant J. 2025 May;122(4):e70234. doi: 10.1111/tpj.70234. Plant J. 2025. PMID: 40437688 Review.
Cited by
-
Molecular Control of Flower Colour Change in Angiosperms.Plants (Basel). 2025 Jul 15;14(14):2185. doi: 10.3390/plants14142185. Plants (Basel). 2025. PMID: 40733424 Free PMC article. Review.
-
Poplar Bud (Populus) Extraction and Chinese Propolis Counteract Oxidative Stress in Caenorhabditis elegans via Insulin/IGF-1 Signaling Pathway.Antioxidants (Basel). 2024 Jul 18;13(7):860. doi: 10.3390/antiox13070860. Antioxidants (Basel). 2024. PMID: 39061928 Free PMC article.
-
Drivers of flower visit and resource sharing between the honeybee and native bees in Neotropical coastal sand dunes.Naturwissenschaften. 2024 Jan 15;111(1):2. doi: 10.1007/s00114-024-01888-7. Naturwissenschaften. 2024. PMID: 38224365
-
Pacific Northwest native plants and native cultivars, part I: pollinator visitation.Environ Entomol. 2025 Feb 19;54(1):199-214. doi: 10.1093/ee/nvae126. Environ Entomol. 2025. PMID: 39800849 Free PMC article.
-
Foraging bee species differentially prioritize quantity and quality of floral rewards.PNAS Nexus. 2024 Oct 7;3(10):pgae443. doi: 10.1093/pnasnexus/pgae443. eCollection 2024 Oct. PNAS Nexus. 2024. PMID: 39411085 Free PMC article.
References
-
- Amsalem E, Grozinger CM, Padilla M, Hefetz A.. 2015. The physiological and genomic bases of bumble bee social behaviour. Advances in Insect Physiology 48: 37–93.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous
