

Corticotropin Releasing Factor Mediates KCa3.1 Inhibition, Hyperexcitability, and Seizures in Acquired Epilepsy
- PMID: 35732494
- PMCID: PMC9337610
- DOI: 10.1523/JNEUROSCI.2475-21.2022
Corticotropin Releasing Factor Mediates KCa3.1 Inhibition, Hyperexcitability, and Seizures in Acquired Epilepsy
Abstract
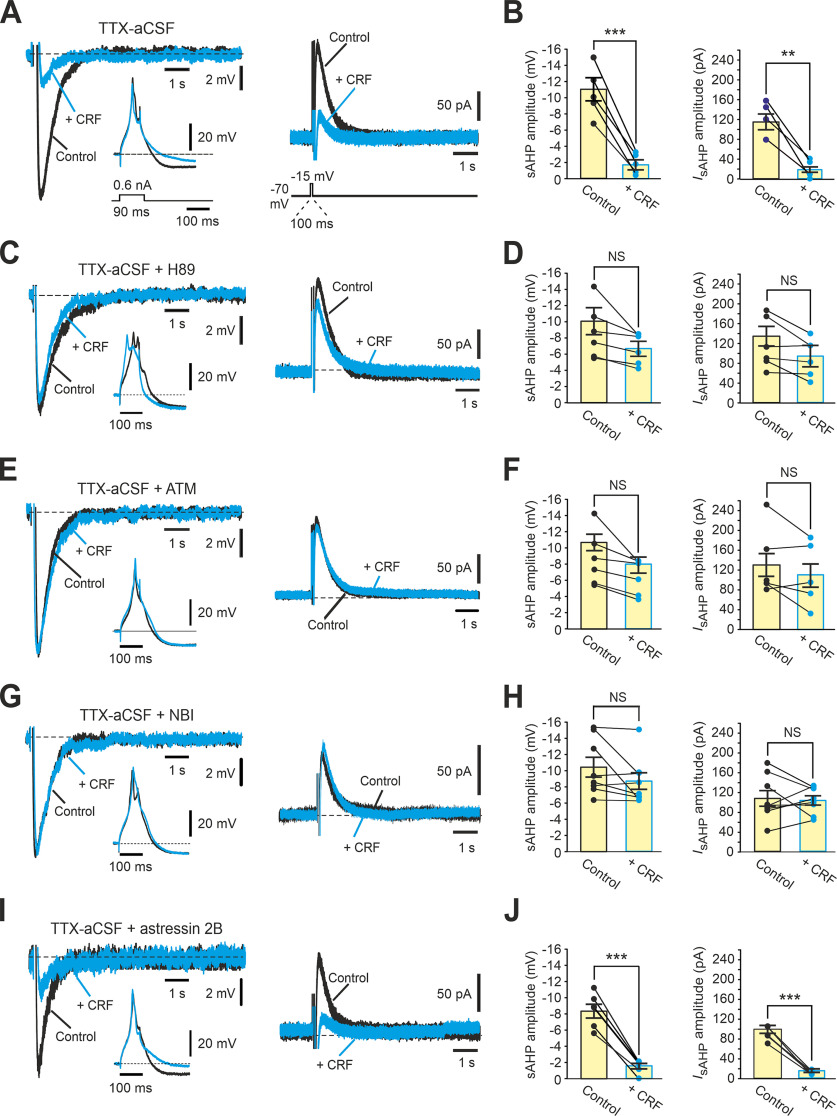
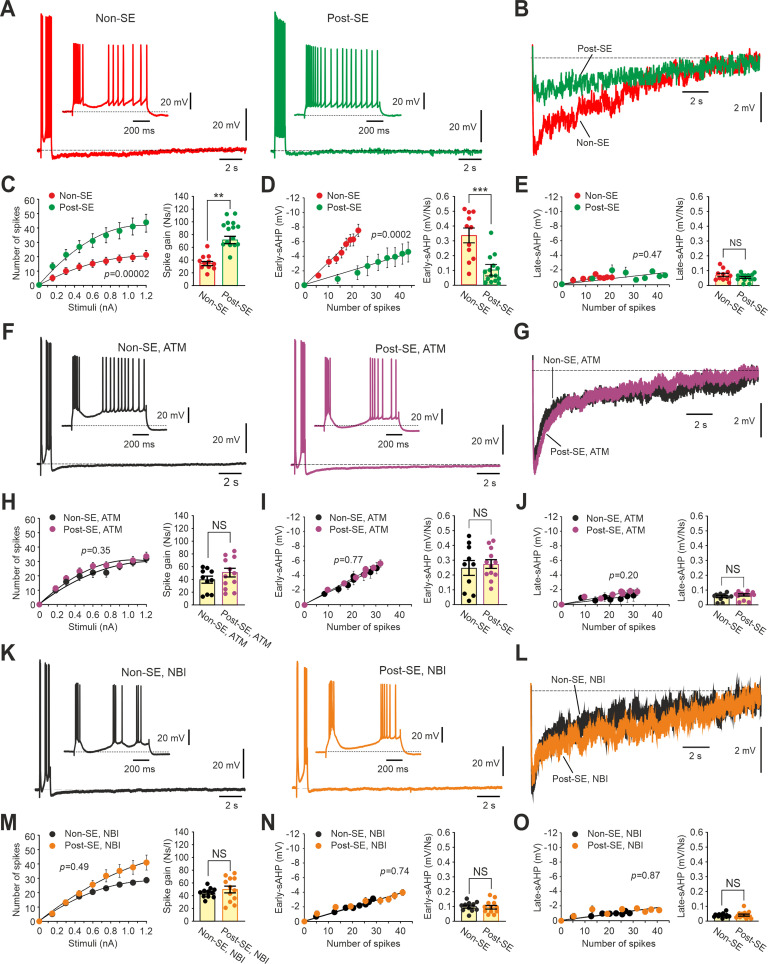
Temporal lobe epilepsy (TLE), the most common focal seizure disorder in adults, can be instigated in experimental animals by convulsant-induced status epilepticus (SE). Principal hippocampal neurons from SE-experienced epileptic male rats (post-SE neurons) display markedly augmented spike output compared with neurons from nonepileptic animals (non-SE neurons). This enhanced firing results from a cAMP-dependent protein kinase A-mediated inhibition of KCa3.1, a subclass of Ca2+-gated K+ channels generating the slow afterhyperpolarizing Ca2+-gated K+ current (IsAHP). The inhibition of KCa3.1 in post-SE neurons leads to a marked reduction in amplitude of the IsAHP that evolves during repetitive firing, as well as in amplitude of the associated Ca2+-dependent component of the slow afterhyperpolarization potential (KCa-sAHP). Here we show that KCa3.1 inhibition in post-SE neurons is induced by corticotropin releasing factor (CRF) through its Type 1 receptor (CRF1R). Acute application of CRF1R antagonists restores KCa3.1 activity in post-SE neurons, normalizing KCa-sAHP/IsAHP amplitudes and neuronal spike output, without affecting these variables in non-SE neurons. Moreover, pharmacological antagonism of CRF1Rs in vivo reduces the frequency of spontaneous recurrent seizures in post-SE chronically epileptic rats. These findings may provide a new vista for treating TLE.SIGNIFICANCE STATEMENT Epilepsy, a common neurologic disorder, often develops following a brain insult. Identifying key cellular mechanisms underlying acquired epilepsy is critical for developing effective antiepileptic therapies. In an experimental model of acquired epilepsy, principal hippocampal neurons manifest hyperexcitability because of downregulation of KCa3.1, a subtype of Ca2+-gated K+ ion channels. We show that KCa3.1 downregulation is mediated by corticotropin releasing factor (CRF) acting through its Type 1 receptor (CRF1R). Congruently, acute application of selective CRF1R antagonists restores KCa3.1 channel activity, leading to normalization of neuronal excitability. In the same model, injection of a CRF1R antagonist to epileptic animals markedly decreases the frequency of electrographic seizures. Therefore, targeting CRF1Rs may provide a new strategy in the treatment of acquired epilepsy.
Keywords: CRF; KCa3.1; channelopathy; hippocampus; intrinsic excitability; temporal lobe epilepsy.
Copyright © 2022 the authors.
Figures







References
-
- Basu T, Maguire J, Salpekar JA (2021) Hypothalamic-pituitary-adrenal axis targets for the treatment of epilepsy. Neurosci Lett 746:135618. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous