

Host control and the evolution of cooperation in host microbiomes
- PMID: 35732630
- PMCID: PMC9218092
- DOI: 10.1038/s41467-022-30971-8
Host control and the evolution of cooperation in host microbiomes
Abstract
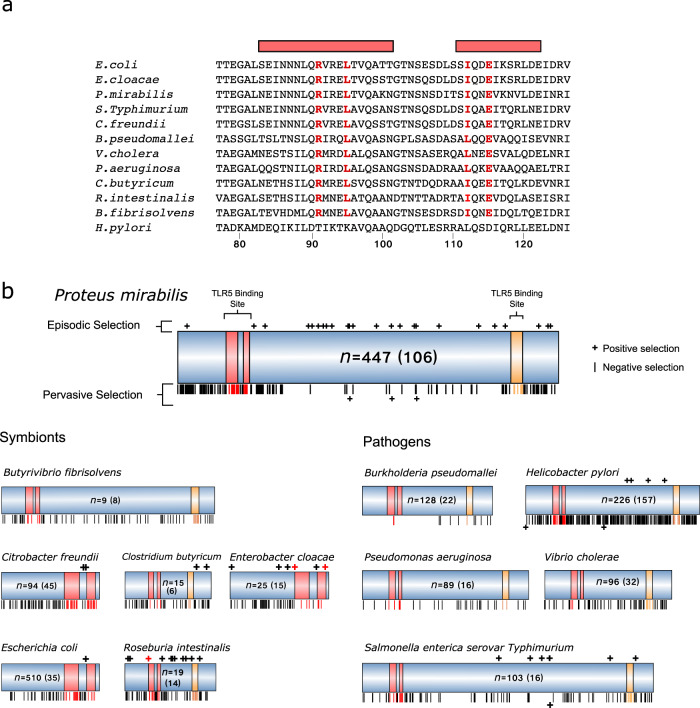
Humans, and many other species, are host to diverse symbionts. It is often suggested that the mutual benefits of host-microbe relationships can alone explain cooperative evolution. Here, we evaluate this hypothesis with evolutionary modelling. Our model predicts that mutual benefits are insufficient to drive cooperation in systems like the human microbiome, because of competition between symbionts. However, cooperation can emerge if hosts can exert control over symbionts, so long as there are constraints that limit symbiont counter evolution. We test our model with genomic data of two bacterial traits monitored by animal immune systems. In both cases, bacteria have evolved as predicted under host control, tending to lose flagella and maintain butyrate production when host-associated. Moreover, an analysis of bacteria that retain flagella supports the evolution of host control, via toll-like receptor 5, which limits symbiont counter evolution. Our work puts host control mechanisms, including the immune system, at the centre of microbiome evolution.
© 2022. The Author(s).
Conflict of interest statement
K.F. is cofounder of Postbiotics plus research LLC.
Figures
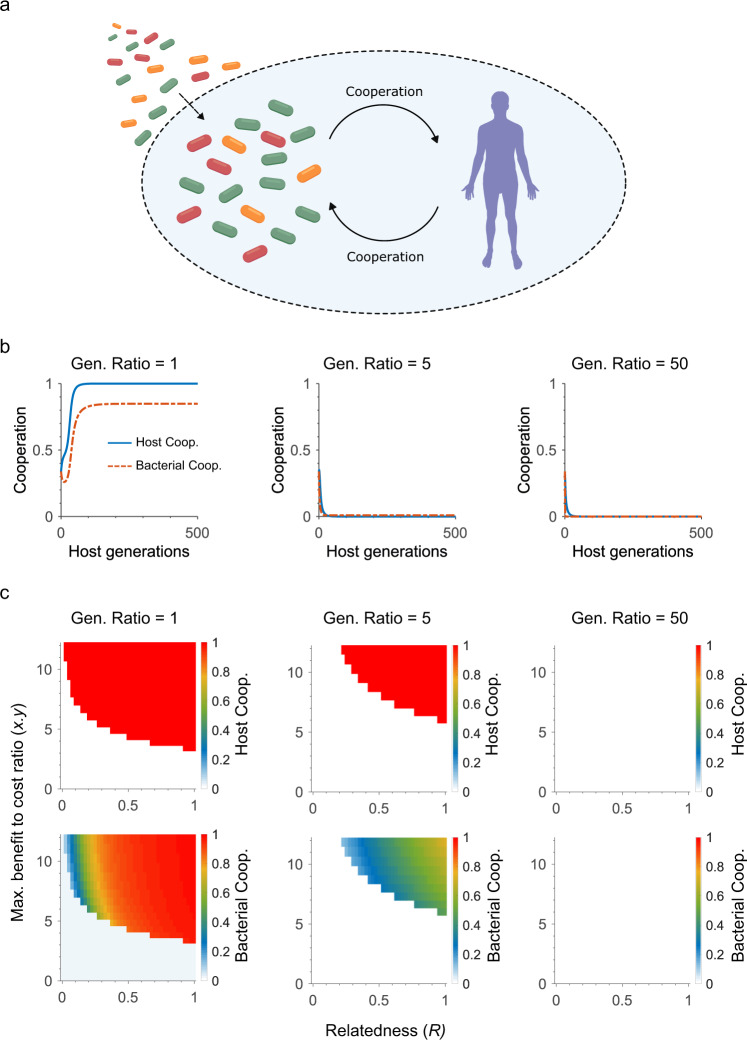
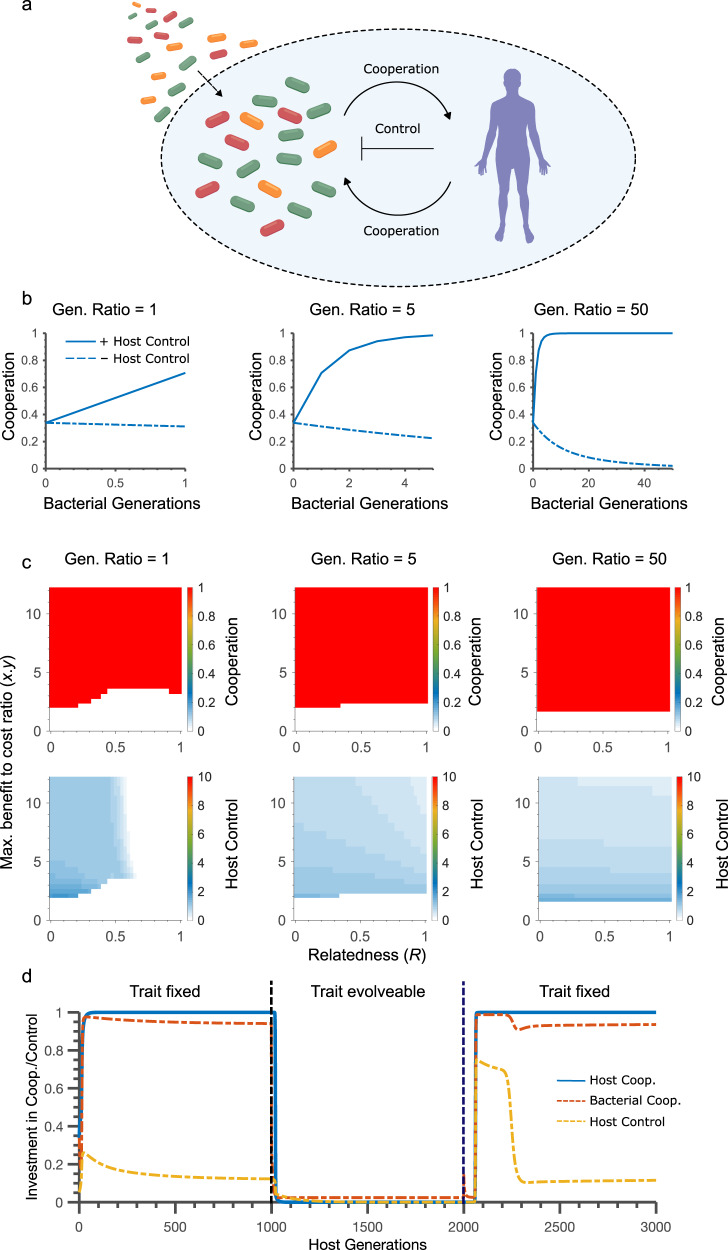
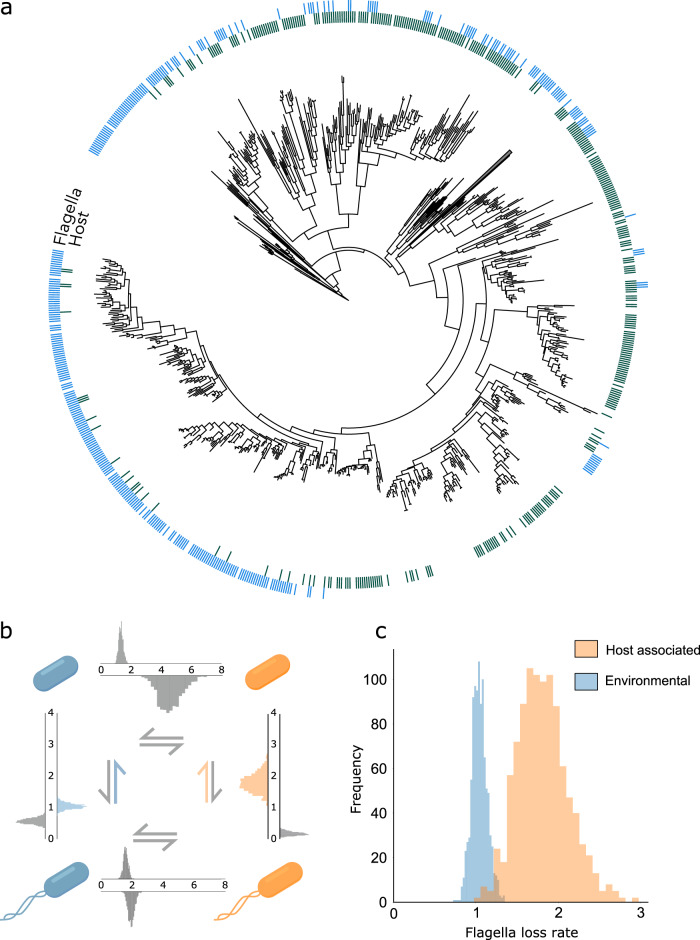
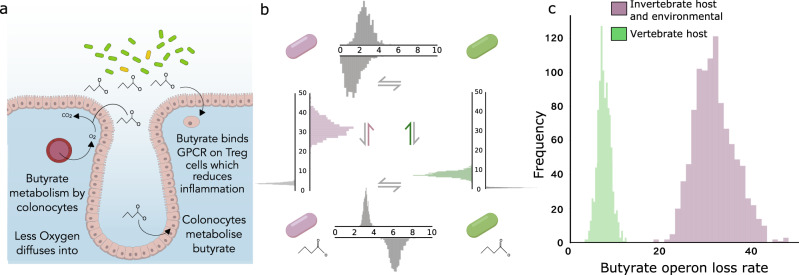
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
