

The evening complex integrates photoperiod signals to control flowering in rice
- PMID: 35733265
- PMCID: PMC9245669
- DOI: 10.1073/pnas.2122582119
The evening complex integrates photoperiod signals to control flowering in rice
Abstract
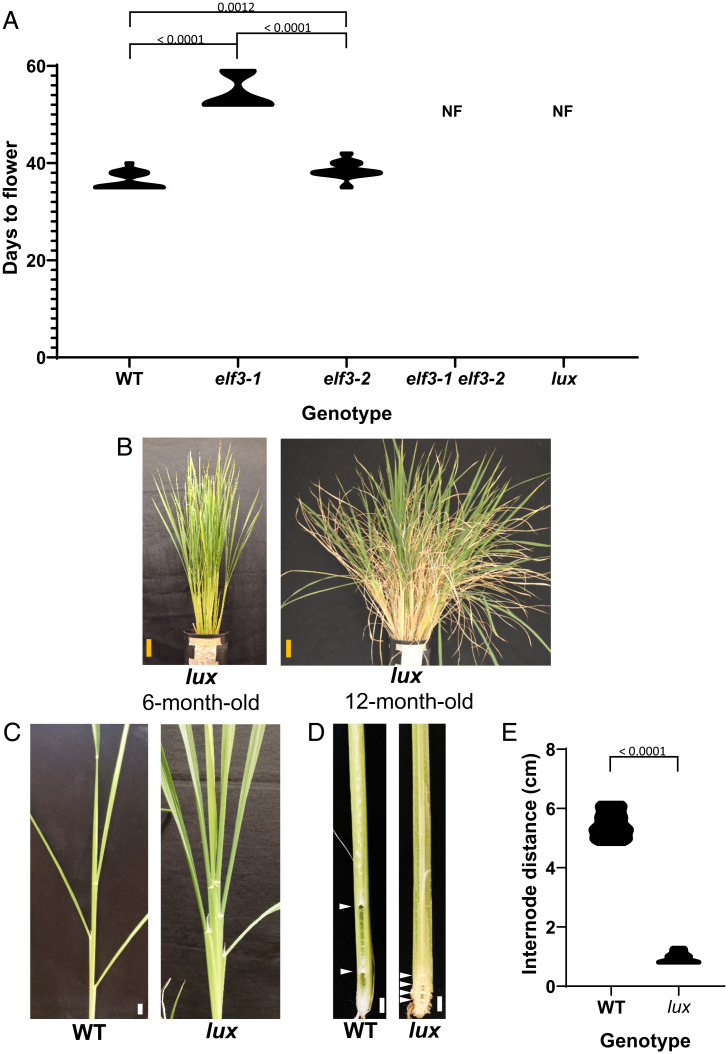
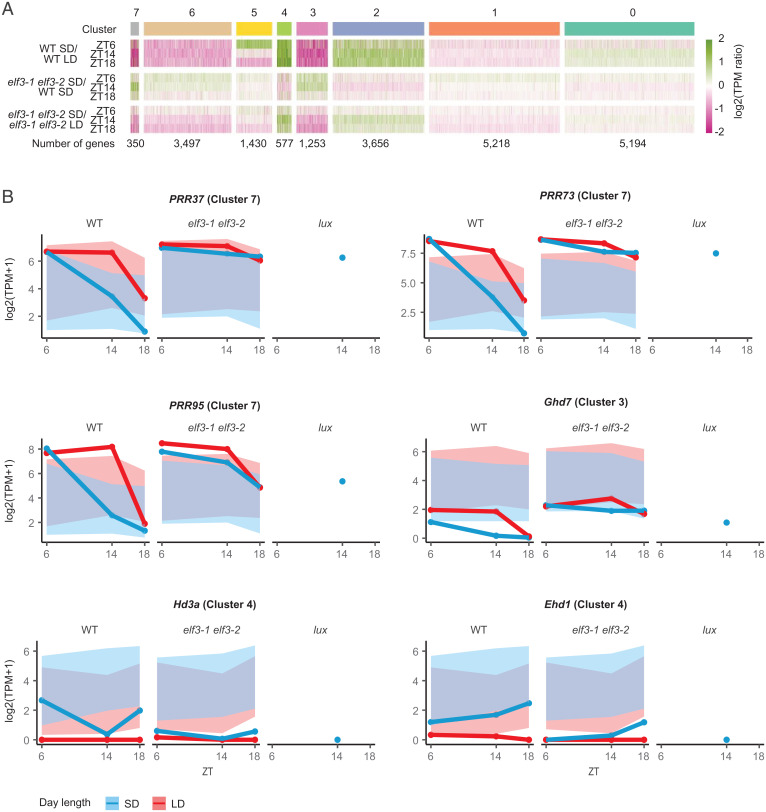
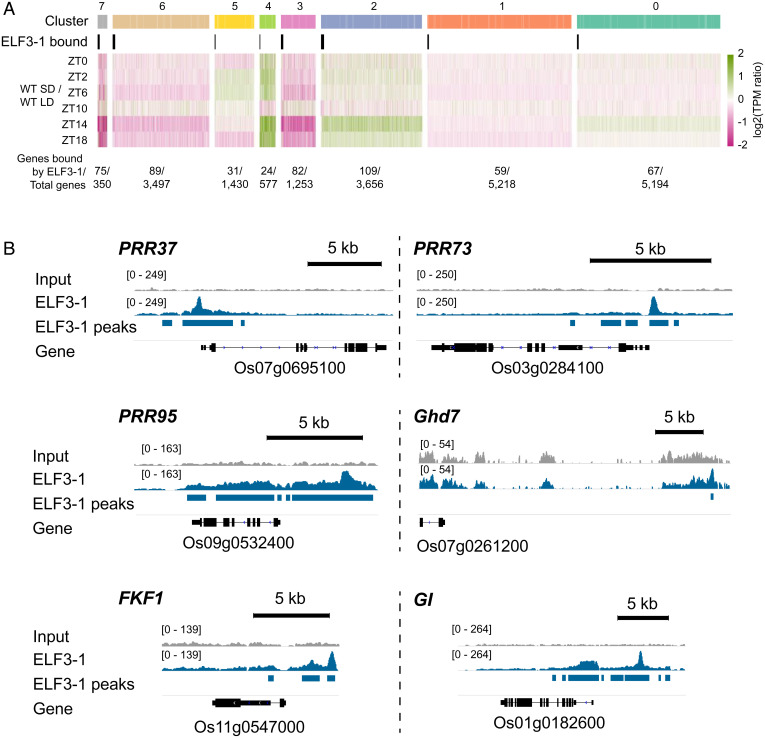
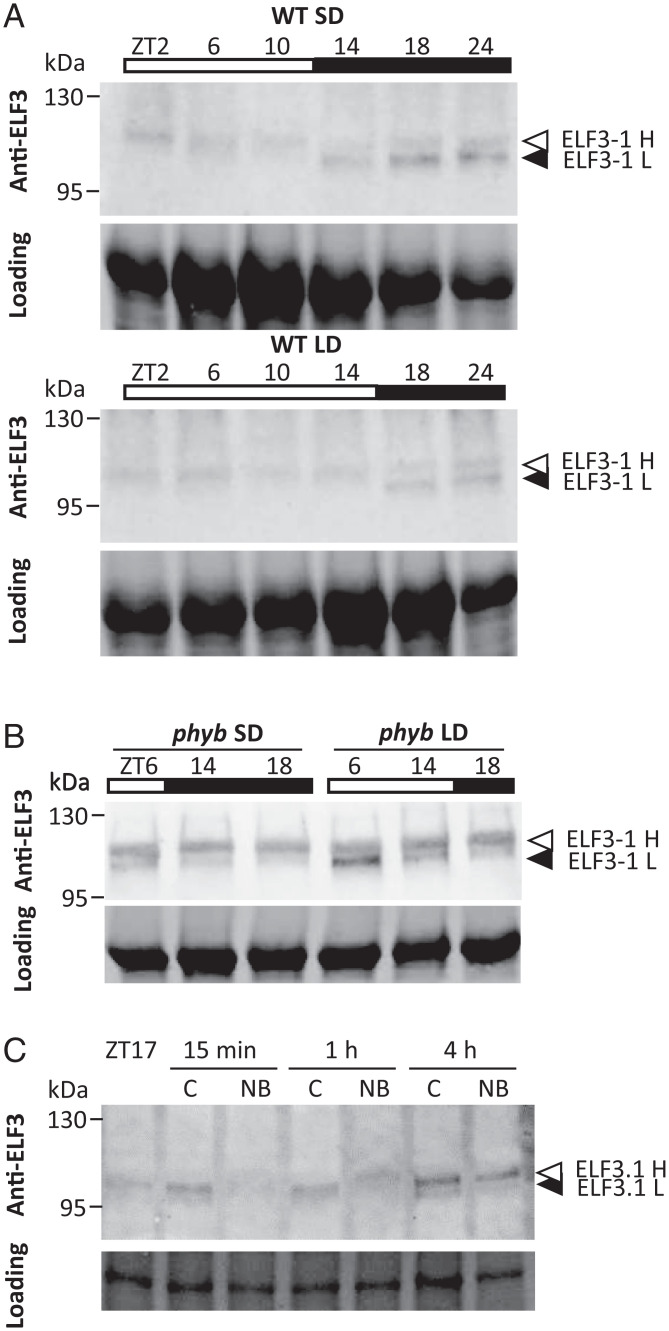
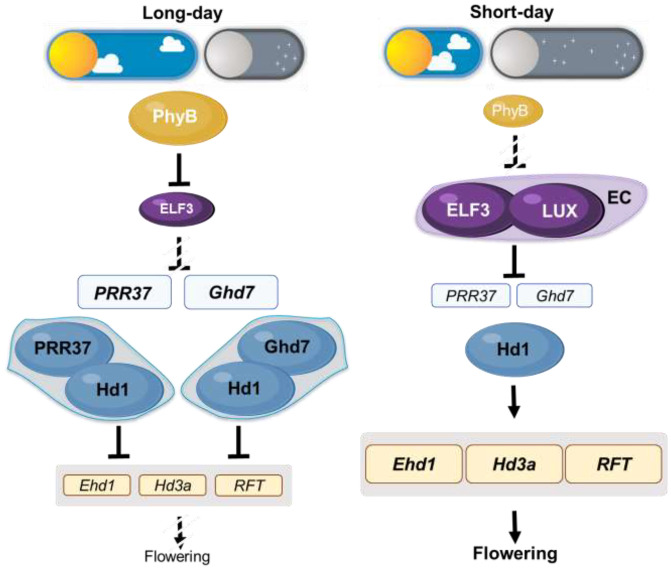
Plants use photoperiodism to activate flowering in response to a particular daylength. In rice, flowering is accelerated in short-day conditions, and even a brief exposure to light during the dark period (night-break) is sufficient to delay flowering. Although many of the genes involved in controlling flowering in rice have been uncovered, how the long- and short-day flowering pathways are integrated, and the mechanism of photoperiod perception is not understood. While many of the signaling components controlling photoperiod-activated flowering are conserved between Arabidopsis and rice, flowering in these two systems is activated by opposite photoperiods. Here we establish that photoperiodism in rice is controlled by the evening complex (EC). We show that mutants in the EC genes LUX ARRYTHMO (LUX) and EARLY FLOWERING3 (ELF3) paralogs abolish rice flowering. We also show that the EC directly binds and suppresses the expression of flowering repressors, including PRR37 and Ghd7. We further demonstrate that light acts via phyB to cause a rapid and sustained posttranslational modification of ELF3-1. Our results suggest a mechanism by which the EC is able to control both long- and short-day flowering pathways.
Keywords: ELF3; Evening Complex; LUX; flowering; rice.
Conflict of interest statement
The authors declare no competing interest.
Figures
Comment in
-
Plants change their clocks to flower at the right time.Proc Natl Acad Sci U S A. 2022 Jul 19;119(29):e2208745119. doi: 10.1073/pnas.2208745119. Epub 2022 Jul 11. Proc Natl Acad Sci U S A. 2022. PMID: 35858367 Free PMC article. No abstract available.
References
-
- Komiya R., Ikegami A., Tamaki S., Yokoi S., Shimamoto K., Hd3a and RFT1 are essential for flowering in rice. Development 135, 767–774 (2008). - PubMed
-
- Kojima S., et al. , Hd3a, a rice ortholog of the Arabidopsis FT gene, promotes transition to flowering downstream of Hd1 under short-day conditions. Plant Cell Physiol. 43, 1096–1105 (2002). - PubMed
-
- Suárez-López P., et al. , CONSTANS mediates between the circadian clock and the control of flowering in Arabidopsis. Nature 410, 1116–1120 (2001). - PubMed
-
- Valverde F., et al. , Photoreceptor regulation of CONSTANS protein in photoperiodic flowering. Science 303, 1003–1006 (2004). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
