

Evaluation of Aspergillus niger Six Constitutive Strong Promoters by Fluorescent-Auxotrophic Selection Coupled with Flow Cytometry: A Case for Citric Acid Production
- PMID: 35736051
- PMCID: PMC9224621
- DOI: 10.3390/jof8060568
Evaluation of Aspergillus niger Six Constitutive Strong Promoters by Fluorescent-Auxotrophic Selection Coupled with Flow Cytometry: A Case for Citric Acid Production
Abstract
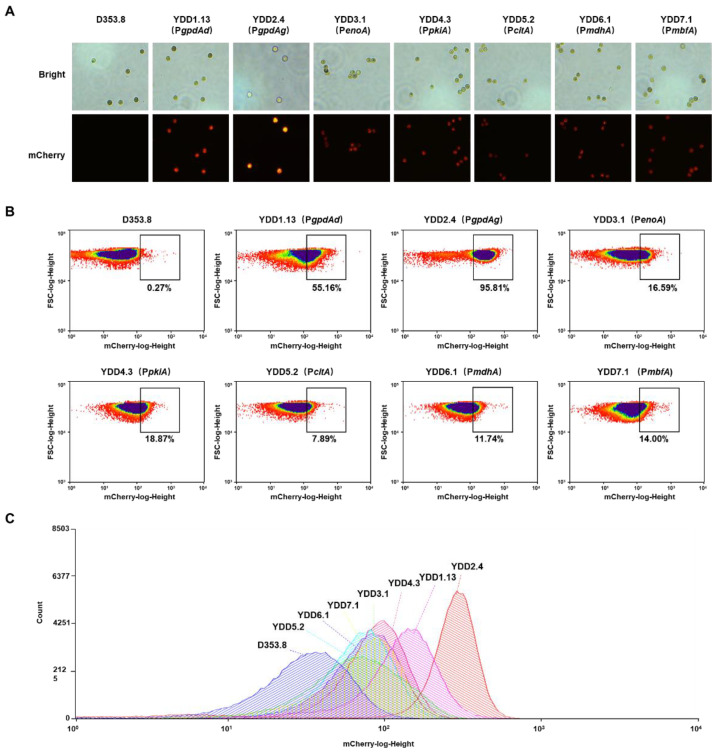
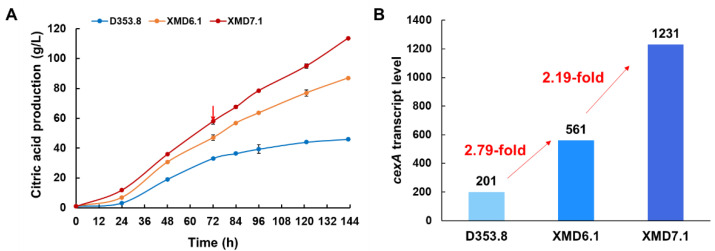
Aspergillus niger is an important industrial workhorse for the biomanufacturing of organic acids, proteins, etc. Well-controlled genetic regulatory elements, including promoters, are vital for strain engineering, but available strong promoters for A. niger are limited. Herein, to efficiently assess promoters, we developed an accurate and intuitive fluorescent-auxotrophic selection workflow based on mCherry, pyrG, CRISPR/Cas9 system, and flow cytometry. With this workflow, we characterized six endogenous constitutive promoters in A. niger. The endogenous glyceraldehyde-3-phosphate dehydrogenase promoter PgpdAg showed a 2.28-fold increase in promoter activity compared with the most frequently used strong promoter PgpdAd from A. nidulans. Six predicted conserved motifs, including the gpdA-box, were verified to be essential for the PgpdAg activity. To demonstrate its application, the promoter PgpdAg was used for enhancing the expression of citrate exporter cexA in a citric acid-producing isolate D353.8. Compared with the cexA controlled by PgpdAd, the transcription level of the cexA gene driven by PgpdAg increased by 2.19-fold, which is consistent with the promoter activity assessment. Moreover, following cexA overexpression, several genes involved in carbohydrate transport and metabolism were synergically upregulated, resulting in up to a 2.48-fold increase in citric acid titer compared with that of the parent strain. This study provides an intuitive workflow to speed up the quantitative evaluation of A. niger promoters and strong constitutive promoters for fungal cell factory construction and strain engineering.
Keywords: Aspergillus niger; CRISPR/Cas9; citric acid; flow cytometry; fluorescence protein; promoter.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



Similar articles
-
Manganese(II) ions suppress the transcription of the citrate exporter encoding gene cexA in Aspergillus niger.Front Bioeng Biotechnol. 2023 Nov 22;11:1292337. doi: 10.3389/fbioe.2023.1292337. eCollection 2023. Front Bioeng Biotechnol. 2023. PMID: 38076415 Free PMC article.
-
Promoter engineering with programmable upstream activating sequences in Aspergillus Niger cell factory.Microb Cell Fact. 2025 Jan 15;24(1):20. doi: 10.1186/s12934-025-02642-y. Microb Cell Fact. 2025. PMID: 39815338 Free PMC article.
-
Engineering of the citrate exporter protein enables high citric acid production in Aspergillus niger.Metab Eng. 2019 Mar;52:224-231. doi: 10.1016/j.ymben.2018.12.004. Epub 2018 Dec 13. Metab Eng. 2019. PMID: 30553933
-
Systems metabolic engineering for citric acid production by Aspergillus niger in the post-genomic era.Microb Cell Fact. 2019 Feb 4;18(1):28. doi: 10.1186/s12934-019-1064-6. Microb Cell Fact. 2019. PMID: 30717739 Free PMC article. Review.
-
Citric acid from Aspergillus niger: a comprehensive overview.Crit Rev Microbiol. 2020 Nov;46(6):727-749. doi: 10.1080/1040841X.2020.1828815. Epub 2020 Oct 12. Crit Rev Microbiol. 2020. PMID: 33044884 Review.
Cited by
-
Genome-wide transcription landscape of citric acid producing Aspergillus niger in response to glucose gradient.Front Bioeng Biotechnol. 2023 Oct 24;11:1282314. doi: 10.3389/fbioe.2023.1282314. eCollection 2023. Front Bioeng Biotechnol. 2023. PMID: 37941722 Free PMC article.
-
Manganese(II) ions suppress the transcription of the citrate exporter encoding gene cexA in Aspergillus niger.Front Bioeng Biotechnol. 2023 Nov 22;11:1292337. doi: 10.3389/fbioe.2023.1292337. eCollection 2023. Front Bioeng Biotechnol. 2023. PMID: 38076415 Free PMC article.
-
A novel sucrose-inducible expression system and its application for production of biomass-degrading enzymes in Aspergillus niger.Biotechnol Biofuels Bioprod. 2023 Feb 13;16(1):23. doi: 10.1186/s13068-023-02274-7. Biotechnol Biofuels Bioprod. 2023. PMID: 36782304 Free PMC article.
-
Exploring and Engineering Novel Strong Promoters for High-Level Protein Expression in Bacillus subtilis DB104 through Transcriptome Analysis.Microorganisms. 2023 Dec 6;11(12):2929. doi: 10.3390/microorganisms11122929. Microorganisms. 2023. PMID: 38138072 Free PMC article.
-
Engineering of Aspergillus niger for efficient production of D-xylitol from L-arabinose.Microb Cell Fact. 2024 Oct 5;23(1):262. doi: 10.1186/s12934-024-02526-7. Microb Cell Fact. 2024. PMID: 39367393 Free PMC article.
References
-
- Meyer V., Cairns T., Barthel L., King R., Kunz P., Schmideder S., Muller H., Briesen H., Dinius A., Krull R. Understanding and controlling filamentous growth of fungal cell factories: Novel tools and opportunities for targeted morphology engineering. Fungal. Biol. Biotechnol. 2021;8:8. doi: 10.1186/s40694-021-00115-6. - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
