

Functional changes of the gastric bypass microbiota reactivate thermogenic adipose tissue and systemic glucose control via intestinal FXR-TGR5 crosstalk in diet-induced obesity
- PMID: 35739571
- PMCID: PMC9229785
- DOI: 10.1186/s40168-022-01264-5
Functional changes of the gastric bypass microbiota reactivate thermogenic adipose tissue and systemic glucose control via intestinal FXR-TGR5 crosstalk in diet-induced obesity
Abstract
Background: Bariatric surgery remains the most effective therapy for adiposity reduction and remission of type 2 diabetes. Although different bariatric procedures associate with pronounced shifts in the gut microbiota, their functional role in the regulation of energetic and metabolic benefits achieved with the surgery are not clear.
Methods: To evaluate the causal as well as the inherent therapeutic character of the surgery-altered gut microbiome in improved energy and metabolic control in diet-induced obesity, an antibiotic cocktail was used to eliminate the gut microbiota in diet-induced obese rats after gastric bypass surgery, and gastric bypass-shaped gut microbiota was transplanted into obese littermates. Thorough metabolic profiling was combined with omics technologies on samples collected from cecum and plasma to identify adaptions in gut microbiota-host signaling, which control improved energy balance and metabolic profile after surgery.
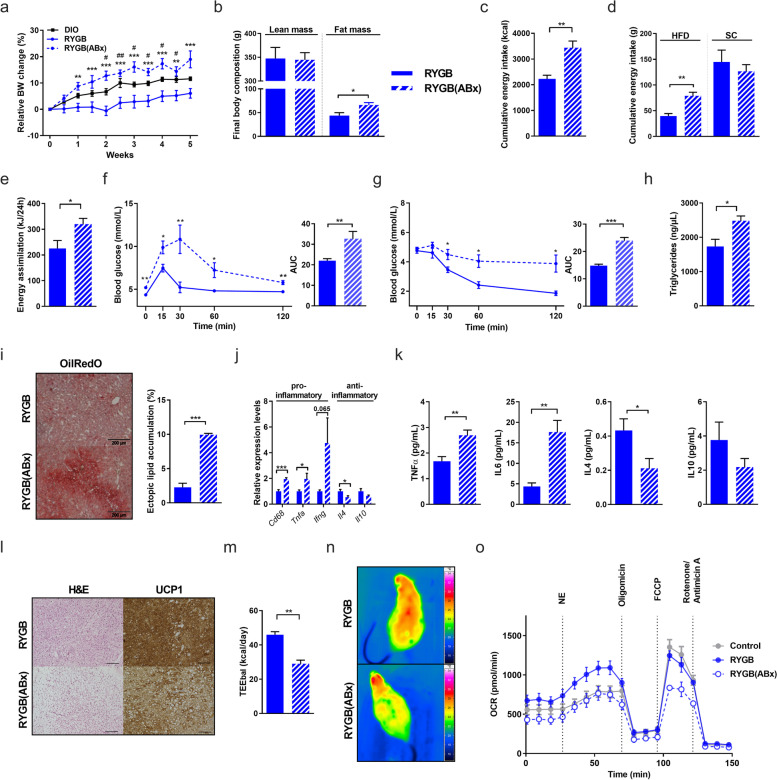
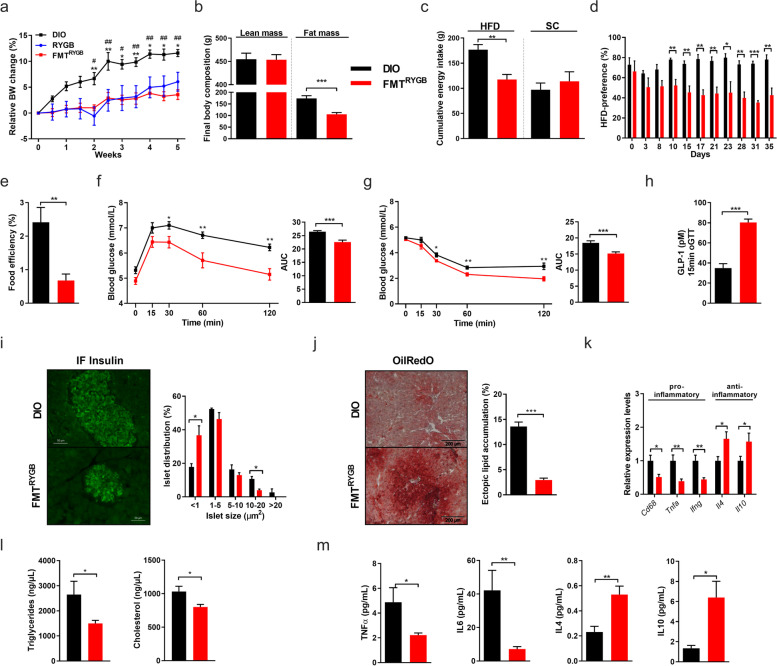
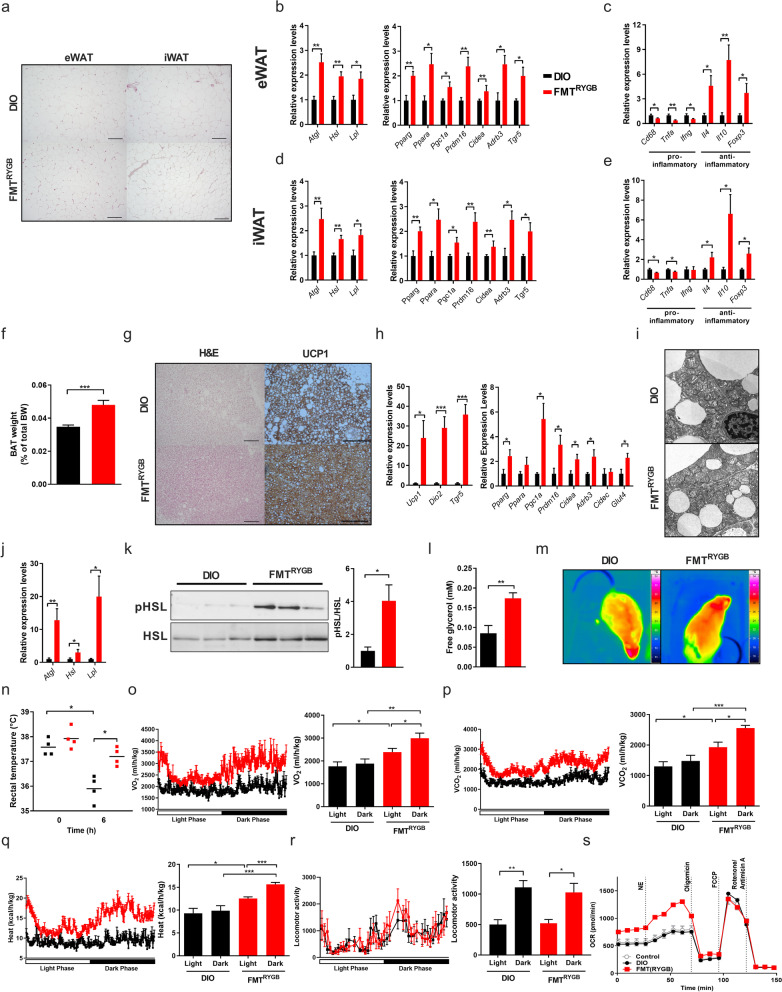
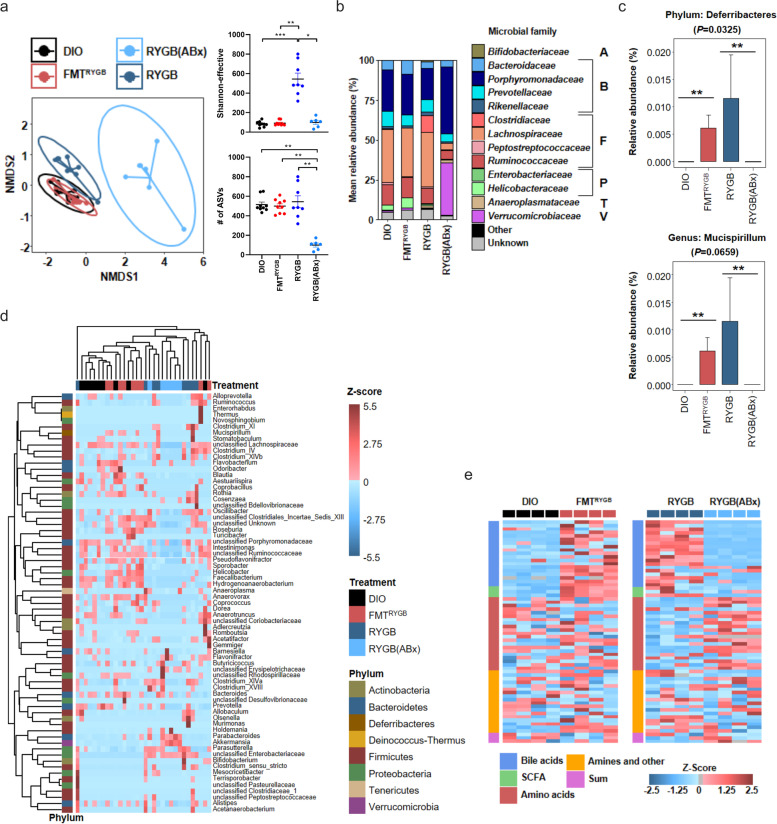
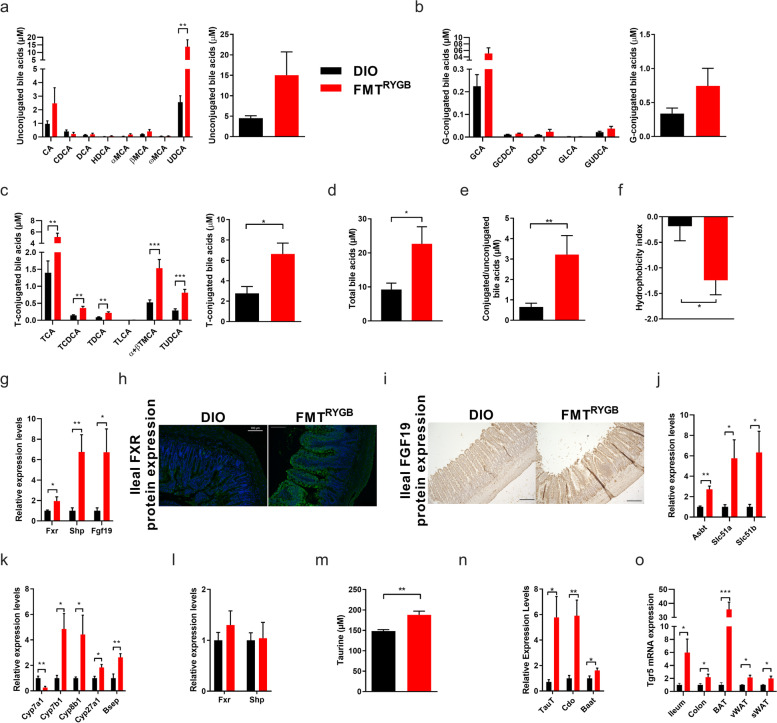
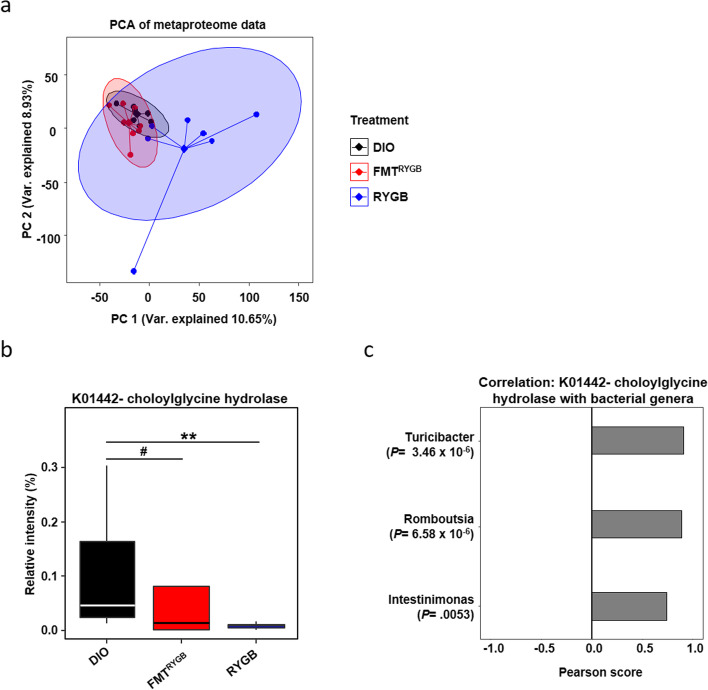
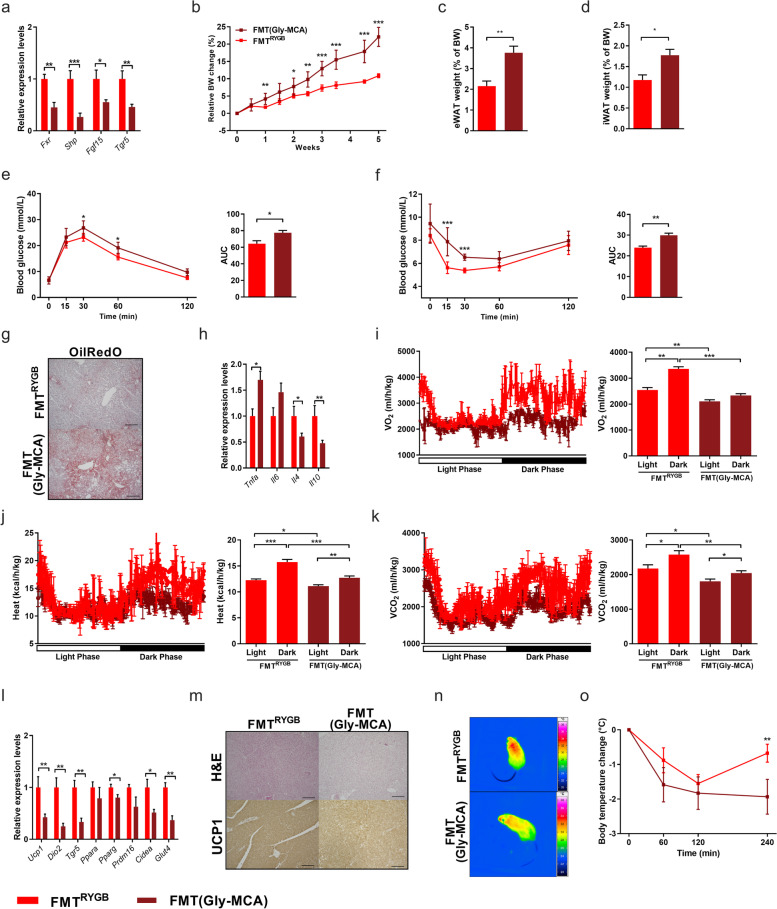
Results: In this study, we first demonstrate that depletion of the gut microbiota largely reversed the beneficial effects of gastric bypass surgery on negative energy balance and improved glucolipid metabolism. Further, we show that the gastric bypass-shaped gut microbiota reduces adiposity in diet-induced obese recipients by re-activating energy expenditure from metabolic active brown adipose tissue. These beneficial effects were linked to improved glucose homeostasis, lipid control, and improved fatty liver disease. Mechanistically, these effects were triggered by modulation of taurine metabolism by the gastric bypass gut microbiota, fostering an increased abundance of intestinal and circulating taurine-conjugated bile acid species. In turn, these bile acids activated gut-restricted FXR and systemic TGR5 signaling to stimulate adaptive thermogenesis.
Conclusion: Our results establish the role of the gut microbiome in the weight loss and metabolic success of gastric bypass surgery. We here identify a signaling cascade that entails altered bile acid receptor signaling resulting from a collective, hitherto undescribed change in the metabolic activity of a cluster of bacteria, thereby readjusting energy imbalance and metabolic disease in the obese host. These findings strengthen the rationale for microbiota-targeted strategies to improve and refine current therapies of obesity and metabolic syndrome. Video Abstract Bariatric Surgery (i.e. RYGB) or the repeated fecal microbiota transfer (FMT) from RYGB donors into DIO (diet-induced obesity) animals induces shifts in the intestinal microbiome, an effect that can be impaired by oral application of antibiotics (ABx). Our current study shows that RYGB-dependent alterations in the intestinal microbiome result in an increase in the luminal and systemic pool of Taurine-conjugated Bile acids (TCBAs) by various cellular mechanisms acting in the intestine and the liver. TCBAs induce signaling via two different receptors, farnesoid X receptor (FXR, specifically in the intestines) and the G-protein-coupled bile acid receptor TGR5 (systemically), finally resulting in metabolic improvement and advanced weight management. BSH, bile salt hydrolase; BAT brown adipose tissue.
Keywords: Bile acids; FXR; Gastric bypass; Gut microbiota; TGR5; Taurine metabolism.
© 2022. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
