

The Good and the Bad: Monocytes' and Macrophages' Diverse Functions in Inflammation
- PMID: 35741108
- PMCID: PMC9222172
- DOI: 10.3390/cells11121979
The Good and the Bad: Monocytes' and Macrophages' Diverse Functions in Inflammation
Abstract
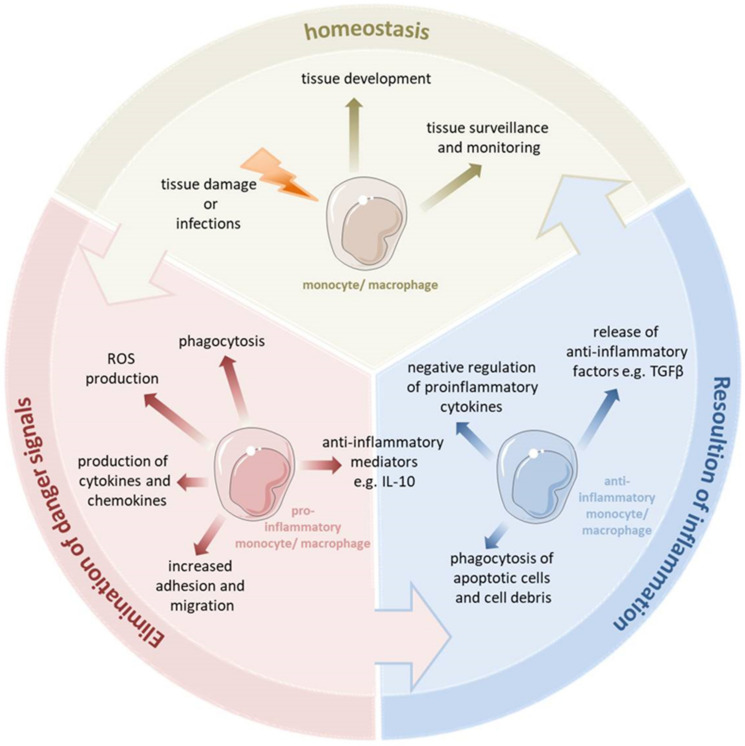
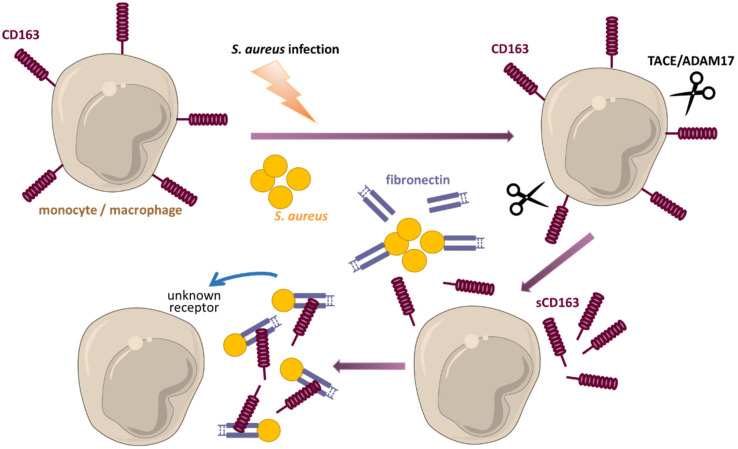
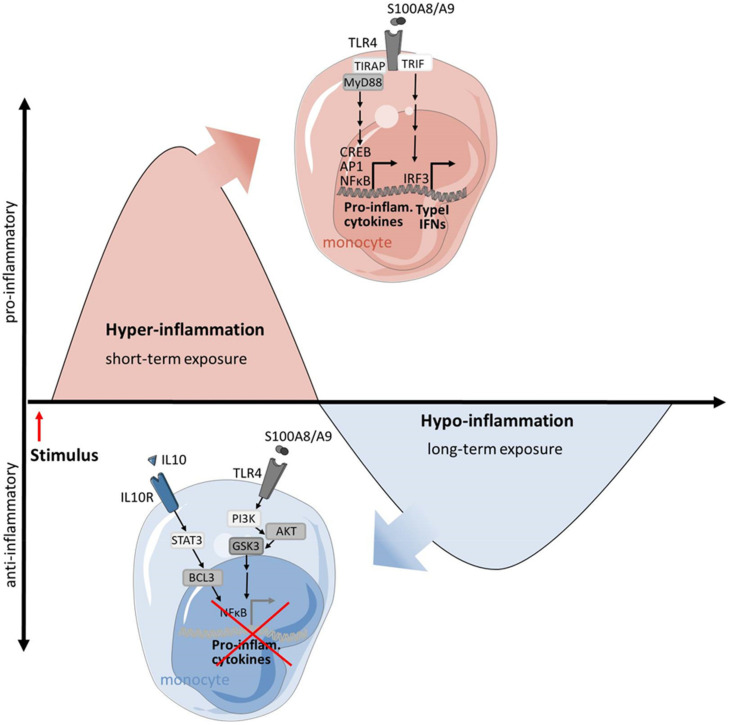
Monocytes and macrophages are central players of the innate immune response and play a pivotal role in the regulation of inflammation. Thereby, they actively participate in all phases of the immune response, from initiating inflammation and triggering the adaptive immune response, through to the clearance of cell debris and resolution of inflammation. In this review, we described the mechanisms of monocyte and macrophage adaptation to rapidly changing microenvironmental conditions and discussed different forms of macrophage polarization depending on the environmental cues or pathophysiological condition. Therefore, special focus was placed on the tight regulation of the pro- and anti-inflammatory immune response, and the diverse functions of S100A8/S100A9 proteins and the scavenger receptor CD163 were highlighted, respectively. We paid special attention to the function of pro- and anti-inflammatory macrophages under pathological conditions.
Keywords: CD163; COVID-19; S100A8; S100A9; SIRS; chronic inflammation; inflammation; macrophage plasticity; macrophages; monocytes.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
